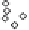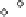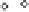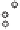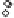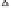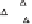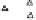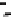Agriculture Reference
In-Depth Information
opportunities available to overcome these con-
straints. Examples of these will be provided
throughout this chapter.
As described earlier, the water demand-supply
balance during the season impacts both biomass
productivity and the partitioning of biomass to set
and fi ll grains. Hence, it is helpful to attempt to
independently characterize this water balance and
use the information during the evaluation of plant
breeding trials. Saulescu and Kronstad (1995)
proposed that a “simulated entry” (i.e., yield esti-
mated using weather and soil data in a dynamic
crop simulation model) be calculated for each
wheat trial and used as an unbiased check to
account for environmental effects. Voltas et al.
(1999b) used environmental indices to explain
genotype × environment interaction in barley
(
Hordeum vulgare
L.) evaluation trials, but these
indices were based on average values of climate
variables during different crop stages and were
not integrative of the water supply-demand
balance. For sorghum (
Sorghum bicolor
), Chapman
et al. (2000) showed that a water demand-supply
index estimated from a crop simulation model
could explain consistent differences in genetic
correlations of line performance between areas
of the sorghum cropping region of northern
Australia.
6
China Loess Plateau
Mediterranean basin
◊
N. American Great Plains
Δ
Southern Australia
5
4
3
2
1
0
0
100
200
300
400
500
600
Evapotranspiration (mm)
Fig. 11.2
Plot of wheat yield and seasonal evapotranspira-
tion for 691 published studies representing four main global
dryland cropping regions. The solid line represents the French
and Schultz (1984) frontier and depicts water-limited grain
yield (
ca.
22 kg ha
−1
mm
−1
). Deviations from the slope refl ect
agronomic, phenology (matching demand for water with
supply), and other factors limiting productivity (Sadras and
Angus 2006).
radiation, act to decrease the realized yield water-
use effi ciency (WUE). The intercept of approxi-
mately 60 mm on the ET axis of Fig. 11.2 is
considered to represent water lost through soil
evaporation or runoff (or drainage), and increases
for locations where numerous small rainfall
events occur within-season (e.g., Mediterranean
environments) rather than prior to sowing. An
increased frequency of such events can contribute
to large water losses through soil evaporation
(e.g., estimated to be in excess of 110 mm by
French and Schultz 1984).
Agronomic factors can explain why the water-
limited yield potential is rarely achieved (French
and Schultz 1984; Sadras and Angus 2006; Sadras
and Rodriguez 2007). These include restricted
water capture (e.g., factors infl uencing root
growth such as late sowing, poor crop establish-
ment, root disease, soil salinity, and acidifi cation),
competition with weeds, nonoptimal sowing
density or arrangement, and leaf disease. In some
cases, the rainfall distribution over the season is
simply out-of-phase with periods of high water
demand in the crop, and there is no opportunity
for the crop to use this water. However, for many
of these water-limiting factors, there are genetic
CHARACTERIZING TARGET
ENVIRONMENTS
Selection for improved performance under
drought is challenging, owing to changes in line
ranking (i.e., genotype × environment interac-
tion) across environments (Ceccarelli 1994). To
enhance efforts in genetic improvement of wheat
yield, it has been argued that it is useful to broadly
defi ne rainfed environment types around timing
and average rainfall amount (Chapman et al.,
2000). An example of such environmental charac-
terization is described by Chapman (2008) for
wheat grown in the northern wheat-growing
region of Australia. There spring wheat is sown
in autumn and grows largely on stored soil water
from monsoonal rains. Mean farm yields are 1.5
to 2.5 t ha
−1
and cropping practices are aimed at