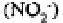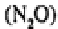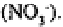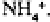Agriculture Reference
In-Depth Information
even higher in seeds. Strong ontogenetic changes occur with much reduced nitrogen
concentrations characteristic of older tissues and minimal concentrations in litter.
In most environments‚ tissue nitrogen concentrations are higher in those species that
have nitrogen-fixing symbionts associated with their roots than in those not so endowed.
As estimated from laboratory cultures‚ mean tissue nitrogen concentrations of ten
species of soil bacteria were 7.7 % while those of fourteen species of soil fungi ranged
from 4.4 to 4.75 %‚ all on a dry weight basis (Anderson and Domsch‚ 1980). Values in
nature may be much lower and Foster (1994) considers that most soil micro-organisms
are in a constant state of near starvation. N concentrations in the fruiting bodies of
a number of field-collected fungi ranged from 1.5 to 7 % of dry weight while mycelial
values as low as 0.68 have been reported from wood-rotting Basidiomycetes (Cowling
and Merrill‚ 1966). Whole termites have N concentrations of the order of 3.5-12.3
%
of
dry biomass (Matsumoto‚ 1976; Spain‚ unpublished) and ants 9.2-12.7 % of dry biomass
(Spain‚ unpublished). In man‚ nitrogen concentrations in soft tissues are approximately
7.2 % while bone contains 4.3 %; hair and nail tissues are somewhat higher at 15-16 %‚
again on a dry weight basis (Bowen‚ 1979).
Considerable energy is required to break the bond in dinitrogen gas‚ whether
this is carried out by organisms‚ or in the production of nitrogenous fertilisers.
Thus‚ the large pool of atmospheric nitrogen is largely unavailable directly to higher
plants and can only be obtained through fixation by lightning and the ability of certain
symbiotic‚ associative and free-living prokaryotes to fix it into useable forms. It is also
unavailable directly to most animals. However‚ N-fixing organisms exist in the intestinal
tracts of many termites (Slaytor and Chappell‚ 1994) (Chapter III.4.3.2.1) and certain
other insects and some evidence exists for N-fixation in the gut of certain earthworms
(Barois
et al
.‚ 1987). The advantages of these symbioses to the host animal are clear given
the very high C:N ratios of the diets of such animals as termites.
Despite the fixation of atmospheric nitrogen by prokaryote organisms‚ most of
the nitrogen required for plant growth in terrestrial ecosystems must be supplied through
faunally‚ microbially - and abiotically - mediated decomposition processes (Amundson
and Davidson‚ 1990)‚ or through fertiliser application. The nitrogen in plant tissues
occurs principally as protein. On tissue death‚ these proteins are initially broken down
through proteolysis and mineralised to yield ammonium ions. These ions may be
adsorbed onto the internal or external surfaces of clays‚ assimilated by plants and
micro-organisms or volatilised after conversion to ammonia gas. The rest largely
undergoes nitrification by aerobic chemautotrophs (or‚ under certain conditions‚ by
heterotrophs) to produce nitrite ions which may be reduced to nitrous oxide
or oxidised further to nitrate ions The nitrate thus formed may move out
of the soil system‚ be taken up by plants or micro-organisms or broken down through
anaerobic denitrification processes (biotic but also abiotic in acid environments) to give
gaseous oxides of nitrogen and dinitrogen gas (Figure I.34).
Higher plants and many micro-organisms may take up inorganic nitrogen either
as or as although micro-organisms preferentially take up inorganic nitrogen
as Certain groups of higher plants may favour a specific form‚ while others appear
to be indifferent. There is evidence that amino acids may form an important source of
nitrogen for plants which may therefore compete with soil micro-organisms (Chapin