Biology Reference
In-Depth Information
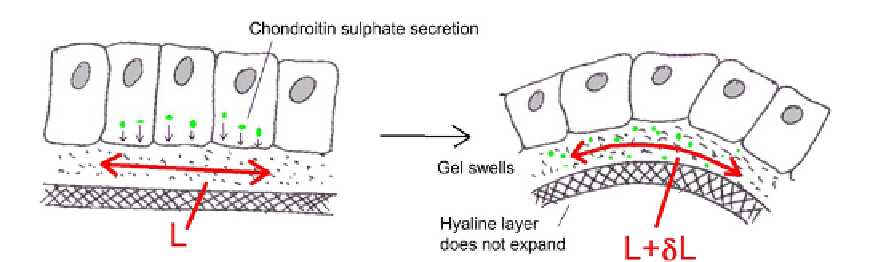
FIGURE 18.6
The gel swelling model for sea urchin invagination. Secretion and swelling of a chondtroitin
sulphate gel expands the apical matrix without expanding the hyaline layer. The extra length, marked
d
L in the
diagram, forces the combination to bend in much the same way that the bimetallic strip in a thermostat is forced to
bend by the unequal expansion of its two layers.
invagination. Embryos lacking RhoA cannot initiate invagination, and precocious activation
of RhoA drives matrix changes normally associated with invagination, and brings that
process forward.
9
This may reflect the activity of Rho in driving secretion of actin-coated
secretory vesicles
14
rather than its activity in driving apical constriction, and illustrates the
difficulties in interpreting inhibition and knockout experiments that concern molecules
that regulate many different aspects of cell behaviour.
MODELS FOR AXIAL INVAGINATION (3):
AN ATTEMPTED SYNTHESIS
It is possible that two entirely different mechanisms of invagination really do exist. It is
also possible that most examples use apical constriction and matrix mechanics together,
but that different experiments are biased towards detecting one or the other. The invagin-
ation of the lens of the eye, which has already been mentioned in
Section II
(Cell Shape
and Cell Morphogenesis) hints at a possible synthesis.
The lens develops from the surface ectoderm that overlies the optic vesicle (the part of the
brain that will form the back of the eye). This ectoderm first thickens to form the lens placode,
an area of thicker epithelium with an elevated cell density compared with the general ecto-
derm next to it. This placode then invaginates to form a sunken bowl, the outer edges of
which fold over and meet. Underneath, the optic vesicle itself invaginates to make the
concave surface of the retina. The effect of the two processes happening together is therefore
invagination in tandem (
Figure 18.7
).
Full invagination of the lens placode involves, and requires, microfilament-mediated
apical constriction. Inhibition of the pathway in
Figure 18.4
at any of a variety of points
d
Shroom3 Trio, RhoA, and ROCK
d
results in only a weak invagination response.
5,8,9
The inhib-
ition of invagination does not, however, block differentiation, which still proceeds and results
in the synthesis of lens crystallins in what is still morphologically surface ectoderm.
5
On the
basis of this evidence alone, it would therefore seem that lens invagination is driven simply
by apical constriction.