Biology Reference
In-Depth Information
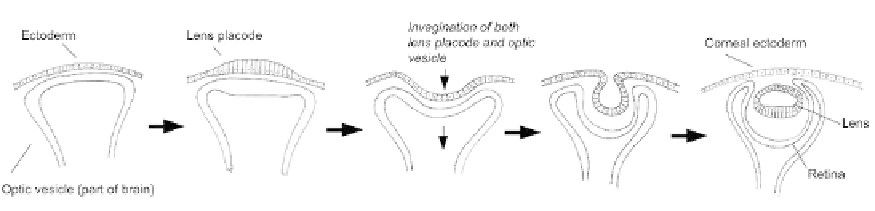
FIGURE 18.7
The double-invagination that takes place in eye development. At the start of the process, the optic
vesicle, an outgrowth of the neural tube, lies just below the surface ectoderm of the head. Part of this ectoderm
thickens by cell elongation to form the lens placode, and this placode begins to invaginate. At the same time, the
underlying optic vesicle itself invaginates. The invagination of the optic vesicle is simple and forms a bowl-shaped
retina. The invagination of the lens is more extreme, so that the 'lips' of the pit meet. Here, neighbour exchange
takes place, so that the lens closes off and drops below the closed-over ectoderm.
The basal side of the lens placode is lined by a prominent extracellular matrix, rich in the
matrix molecule fibronectin. Ablation of the fibronectin gene a few hours before invagination
is due to start (using Tamoxifen-activated cre-mediated recombination) blocks invagination
without affecting parameters such as cell proliferation or early lens differentiation.
15
The
ectoderm even shows intense apical expression of actin and myosin (although it is not clear
whether any contraction takes place). On its own, this evidence would suggest that invagin-
ation of the lens, like that of the sea urchin archenteron, is driven by a matrix-driven mech-
anism rather than a cytoskeletal one.
Taken together, the two lines of evidence clearly indicate that both cytoskeletal and matrix-
based mechanisms are required for lens invagination to take place properly. Detailed exam-
ination of the consequences of fibronectin loss suggests an early failure, at the stage of
making a placode before invagination proper has begun; the density of cells does not rise
and the epithelium fails to thicken, although rates of cell proliferation are still normal. The
mechanisms of placode formation are not fully understood, but one possible contribution
that the matrix may make is to restrain the spread of cells away from the placode area as
they multiply, so that their density has to increase and so that they have to elongate to fit
in, thereby making the thickened placode automatically.
15
This idea will only work if the
speed of proliferation exceeds the cells' ability to remodel the matrix to accommodate it,
but the idea does have support in principle from a 1960s observation that mechanically
restraining the expansion of ectoderm is sufficient to cause placode formation even in a place
where no placode should be.
16
The idea of matrix providing mechanical constraint may also address a logical deficiency
of the apical constriction model. Apical constriction will result in the production of wedge-
shaped cells only if something prevents the basal side of the cell from reducing its diameter,
because, if this were allowed, then the lowest energy response to active apical thickening may
well be production of thin, columnar cells rather than wedge shapes that will deform an
entire epithelial sheet (
Figure 18.8
, upper). If attachment to a thick, relatively unyielding
matrix held the basal area of the cells constant, then basal thinning would be impossible
and wedges would have to result (
Figure 18.8
, lower). In this model, efficient invagination
requires both apical constriction and matrix mechanics.