Biology Reference
In-Depth Information
recruitment of XRCC4 or XLF to DSBs,
124,127
however,
chromatin fractionation studies suggest that DNA-PKcs
is required for recruitment of XRCC4-DNA ligase IV to
chromatin.
199
DNA end processing by PNKP presum-
ably occurs when PNKP is recruited to DSBs by
XRCC4 and/or the XRCC4-DNA ligase IV complex.
171
When other processing events occur is not yet known.
Regardless, once ends have been processed, they are
ligated by DNA ligase IV in a reaction that is regulated
by XRCC4 and XLF (
Figure 8.8
). How and when the
NHEJ proteins are released from DNA once rejoining
is complete is also not understood, although studies in
Xenopus
extracts suggest that ubiquitination may play
a role in removal of Ku from DSB ends.
200
The critical question of how NHEJ occurs in the
context of chromatin is only beginning to be addressed.
In recent studies, the ATM-dependent chromatin
remodeling proteins ACF1 and SNF2 were shown to
be recruited to DSBs and ACF1 was shown to interact
directly with Ku70.
201
Moreover, ablation of the histone
acetyltransferases CBP and p300 suppressed NHEJ sug-
gesting roles for chromatin remodeling and chromatin
relaxation in facilitating NHEJ.
202
The NHEJ Pathway in V(D)J Recombination
In V(D)J recombination, variable (V), diversity (D),
and joining (J) segments in immunoglobulin and T cell
receptor genes are excised and rejoined to create anti-
body diversity. The process is initiated by the recombi-
nase activating genes RAG1 and RAG2 which create
DSBs that are then ligated by components of the classical
NHEJ pathway. DNA-PKcs, Artemis, Ku, XRCC4, XLF
and DNA ligase IVare all essential for V(D)J recombina-
tion. Thus, V(D)J recombination can be regarded as
a specialized form of NHEJ, limited to T and B cells,
that is optimized to produce DNA sequence diversity
at DNA ends. Apart from RAG1, RAG2, and TdT, which
are lymphoid specific, all of the other NHEJ proteins are
widely expressed in mammalian tissues and probably
play similar roles in V(D)J recombination and repair of
IR-induced DSBs. Here we will provide a brief overview
of the process of V(D)J recombination, describing the
role of the NHEJ pathway. The reader is also referred
to the many excellent in-depth reviews for additional
information on this topic.
66,101,102,151,203
In V(D)J recombination, variable (V), diversity (D)
and joining (J) segments in IgG and T cell receptor
(TCR) genes are excised and rejoined to create antibody
diversity. V(D)J recombination is initiated by the RAG1/2
complex which cleaves dsDNA at specific recombina-
tion signal sequences (RSS) that flank gene segments to
be excised and rejoined (
Figure 8.9
). The endonuclease
activity of the RAG1/2 complex results in formation of
a DNA single-strand break with a 3
0
-OH at the V, D or
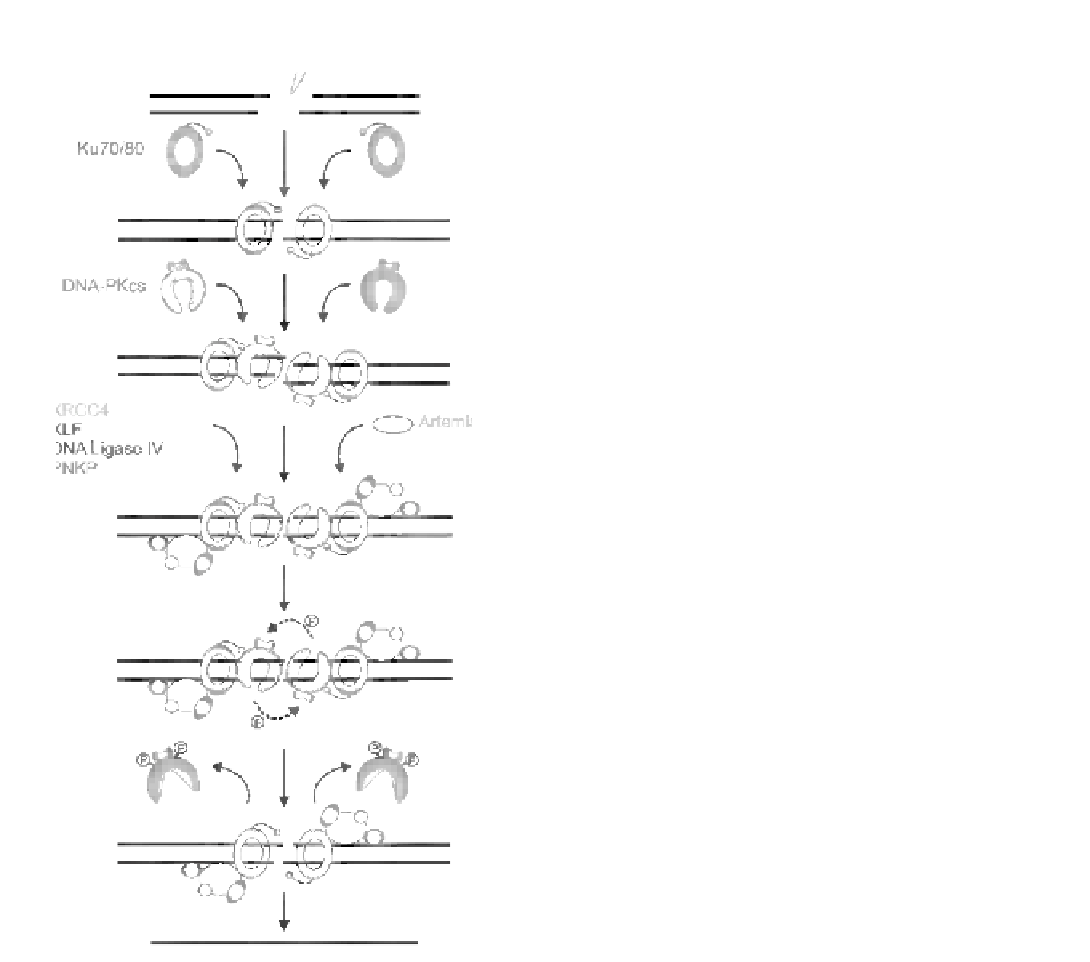
FIGURE 8.8
Model for repair of IR-induced DSBs by NHEJ. IR
induces a complex DSB with non-ligatable ends and other damage
surrounding the DSB. Ku binds to the ends of the DSB and trans-
locates inwards, leading to recruitment of DNA-PKcs (which then
occupies the DSB termini) and stimulation of DNA-PKcs protein
kinase activity. Two DNA-PKcs molecules on opposite sides of the
DSB interact to form a synaptic complex. At an undefined step prior to
ligation, processing factors such as PNKP (which interacts with the
XRCC4-DNA ligase IV complex and/or XRCC4) and Artemis (which
interacts with DNA-PKcs) as well as the XRCC4-DNA ligase IV
complex and XLF (which interacts with XRCC4 and/or XRCC4-DNA
ligase IV) are recruited to the Ku
e
DNA complex. Whether DNA-PKcs
is required for the recruitment of PNKP and XRCC4-DNA ligase IV
and XLF is either not known (PNKP) or unclear (XRCC4-DNA ligase
IV/XLF). DNA-PKcs undergoes autophosphorylation in
trans
, which
causes a conformational change that releases DNA-PKcs from the
DSB. After processing by PNKP, Artemis and other factors, precisely
which ones depending on the forms of damage at the termini and in
the vicinity of the break, the XRCC4-DNA-ligase IV complex rejoins
the DNA ends to repair the DSB. The activity of DNA-ligase IV is
regulated by XLF.
From Dobbs
et al.
, 2010;
162
see text for details.