Biology Reference
In-Depth Information
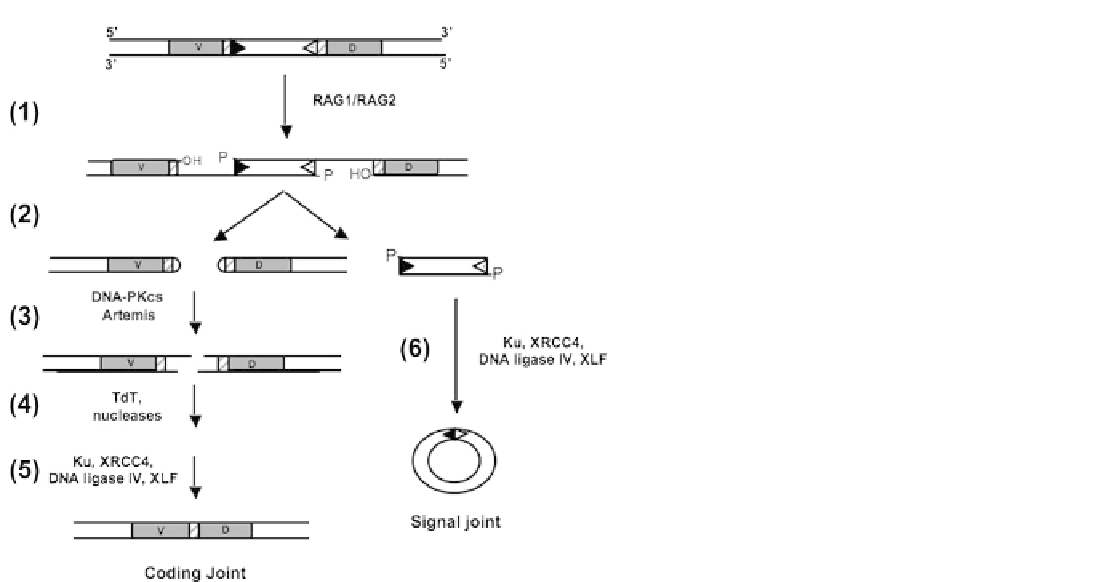
FIGURE 8.9
Model for repair of RAG-induced DSBs by
V(D)J recombination: The gene to be processed contains V
and D segments (coding sequences, gray rectangles) flanked
by recombination signal sequences (RSS, triangles). (1)
Cleavage is initiated by the RAG1/RAG2 endonuclease
complex (modulated by HMG proteins) to create a nick with
a3
0
-OH group and a 5
0
-phosphate group 5
0
to the RSS. (2)
The two RSSs plus the intervening DNA are excised leaving
the two coding ends with closed DNA hairpin ends. (3) The
coding end hairpins are opened by Artemis in complex with
DNA-PKcs in a reaction that requires phosphorylation,
likely of DNA-PKcs. (4) The ends of the opened termini are
processed by nucleases (creating nucleotide loss) and TdT
(which adds nucleotides) to create additional diversity at the
DNA ends. (5) The processed coding ends are ligated by the
NHEJ factors Ku, XRCC4, DNA ligase IV and XLF to create
the coding joint which ultimately is expressed as the coding
sequence. (6) The RSSs are rejoined (signal joint) by the
NHEJ factors Ku, XRCC4, DNA ligase IV and XLF to create
a circular DNA molecule that has no further function and is
lost from the system. Note: recent studies suggest DNA-
PKcs and ATM have redundant roles in signal joint forma-
tion. See text for additional details.
J termini (which will become the coding end) and
a5
0
-phosphate group at the RSS termini (which will
become the signal end). The coding end 3
0
-OH then
attacks the opposite strand in a non-enzymatic reaction,
producing a closed DNA hairpin at the coding end.
Artemis and the catalytic activity of DNA-PKcs are
required for opening of the coding end hairpins, while
Ku, XRCC4, DNA ligase IV, and XLF are required for
rejoining both coding and signal ends (see
Figure 8.9
).
Recent studies also suggest that whereas DNA-PKcs is
absolutely required for coding joint formation, DNA-
PKcs and ATM have overlapping roles in signal joint
formation.
88,204
Moreover, functionally redundant roles
between ATM and XLF have been revealed in repair of
V(D)J substrates in the context of chromatin;
205
but
whether this translates to functional redundancies
between ATM and XLF in repair of IR-induced chro-
matin associated DSBs remains to be determined.
Coding ends are also subject to limited nuclease
digestion prior to rejoining and the lymphoid specific
DNA polymerase X family member, TdT, fills in
missing sequence leading to further diversity at coding
junctions. As in NHEJ of IR-induced DSBs, autophos-
phorylation of DNA-PKcs is important for V(D)J recom-
bination, regulating DNA end processing by protection
from nuclease attack with different phosphorylation
sites have opposing effects on V(D)J rejoining.
151
The
joined coding sequences go on to form the protein
products whereas DNA containing signal joints are
eliminated from the system. As discussed below, misre-
pair by either NHEJ or V(D)J recombination has the
potential to lead to chromosomal aberrations, including
chromosomal translocations.
N
HEJ AND TUMORIGENES
IS
Role of NHEJ in Generating Chromosomal
Aberrations
Carcinogenesis is regarded as a multi-step process
involving several mutations. Mutations can include
point mutations, deletions or insertions of DNA, and
chromosomal rearrangements such as translocations.
In considering the role of NHEJ in the process, it is
important to realize that while a primary function of
this DNA repair pathway is to maintain genome
stability, it is nonetheless an error-prone pathway
because typically between 1 and 20 nucleotides can be
lost from each DNA end at NHEJ-mediated rejoining
events. In addition, NHEJ may cause translocations by
misjoining the ends arising from two or more double-
strand breaks. Thus, it is possible that NHEJ may
contribute to both genome protection and mutation.
On the other hand, despite the fact that HR is intrinsi-
cally an error-free process, it has been pointed out
that, except when sister chromatids are available in
late S and G2 phases of the cell cycle, HR could cause
crossovers through inappropriate alignment of repeti-
tive sequences (e.g. Alu repeat sequences).
206
This
may explain why HR is restricted to the late S and G2
phases.