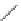Geoscience Reference
In-Depth Information
V
min
= f{
j
}
HIGH
Cell Loses Ability to Divide
Cells Divide
& Grow
V(
j,t
) =
f{
j
, G(
t
)}; G(
t
) = 1
External Growth Rate
G(t)
and position
j
define rate of cellular
division
Cells
unable to
divide
V(
j,t
) =
f{
j
, G(
t
)}; G(
t
) < 1
Cells
differentiate
and mature
V(
j,t
) =
f{
j
, G(
t
)}; G(
t
) << 1
Cells Enter Dormancy
V
cr
0
0
Position (
j
) of cell from cambial initial
Fig. 3.3
Schematic diagram showing the functions that control cell division and transitions in the
Vaganov-Shashkin model. The rate of cambial cell division (which is proportional to the cellular
growth rate
V
(
j
,
t
)) is a linear function of the position (
j
) of the cell in the cellular file and the daily
external environmental growth rate
G
(
t
). The division rate increases with distance from the cambial
initial. The exponential function
V
min
(
j
) defines the threshold rate of division, below which cells
lose the ability to divide and mature to xylem cells. The size of the actively dividing cambial zone
is therefore a nonlinear function of the rate of cellular division. The third function,
V
cr
, defines the
division rate at which cells still in the cambium enter dormancy
with maximum rates of cell division observed tangential to the zone of radial expan-
sion (the 'cambial edge'). Radial tracheid dimension is mainly determined during
cell production and at the beginning of expansion.
The model uses daily precipitation and temperature from meteorological stations
as its required input data. The 28 primary model parameters are based on empiri-
cal and experimental data, whose selection is discussed in detail by Vaganov et al.
gies, simulated growth rates, and number of cambial cells, are solely a function of
those environmental and biological activities modeled in the Growth and Cambial
Blocks. Hence, simulations do not reflect direct growth influences due to increasing
atmospheric CO
2
concentration over the past 150 years. Nor are additional biolog-
ical or ecological influences on patterns of tree-ring formation modeled, including
those caused by tree age or geometry, interseasonal carbon storage, canopy and
root activity, or stand-level competition and disturbance. In a sense the simulations
can be considered 'idealized' mean site tree-ring chronologies with respect to the
modeled processes.