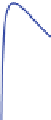Geoscience Reference
In-Depth Information
and so remain for some considerable time, safely below the maximum size by some
margin.
This, of course, is a great simplification of likely tree mortality influences. It takes
no account of the competition dynamics of trees growing within forest stands; i.e.,
dence, particularly when competition pressures are not strong, that to become old
strate this concept using subfossil pine ring width data from northern Fennoscandia
the last 7500 years, but here we have excluded any data from trees that were alive
after 1724. This choice precludes any human sampling influence on the life span
of trees. The retained data were used to produce a single RCS curve. The measure-
ment data for each tree sample were then summed to provide an estimated diameter,
and this diameter was compared to the mean regional diameter calculated from the
appropriate age point of the RCS curve to give a relative growth rate (i.e., the ratio
of tree diameter at its final year to the RCS curve diameter at that year). In this way,
all the individual tree measurement series were grouped into eight relative growth
classes and a sub-RCS curve was constructed for each relative class. The mean
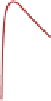
growth rate (millimeters per year) by age for these eight RCS curves is shown in
with age. A tendency for higher growth-rate classes to have shorter life spans can
Fig. 5.5
Based on 1724 subfossil trees from Torneträsk (Grudd et al. 2002) and Finnish Lapland
(Eronen et al. 2002) from the period circa 5400 BC to AD 1724; i.e., excluding trees that were alive
after 1724 to avoid any anthropogenic factors and sampling bias: (
a
) all trees sorted by growth rate
relative to a single regional curve standardization (RCS) curve, into eight separate RCS curves; (
b
)
a scatter plot of mean growth rate for each tree plotted against final tree age. Scatter plot points are
shaded
to match their growth rate curve of (
a
). This figure illustrates the tendency for longevity to
be inversely proportional to tree vigor