Geoscience Reference
In-Depth Information
r
r
q
q
r
r
q
q
r
q
Figure 6.12
Pathways of water vapour and CO
2
out and into a leaf. Both encounter
the boundary-layer resistance
r
b
and the variable stomatal resistance
r
s.
In addition,
CO
2
has to pass a number of cell interfaces, relected by the mesophyll resistance
r
m
.
(After Willmer and Fricker,
1996
)
where
q
i
and
q
e
are the speciic humidity inside the substomatal cavity and just above
the stomate, respectively, and
r
s
is the stomatal resistance (compare the aerodynamic
resistances discussed in
Chapter 3
). Another parameter often used to express the
effect of the stomata is the stomatal conductance
g
s
, which is simply the reciprocal
of the stomatal resistance (
g
s
= 1/
r
s
) which gives
g
s
the units of a velocity. In plant
physiology literature luxes are often given as molar luxes, rather than mass luxes.
Then
g
s
is used with units of mmol m
-2
s
-1
where at the same time the concentration
is given as mole fractions.
The air inside the substomatal cavity is considered to be saturated with water
vapour, and hence
q
i
is equal to the saturated speciic humidity at the temperature of
the leaf:
qq
r
−
()
T
T
=−
ρ
e
sat
s
(6.21)
s
This implies that there is a clear link between transpiration and the temperature of the
leaves. This notion is relevant in the context of the microclimate within the canopy, as
the temperature may vary vertically (see
Section 6.6
). Furthermore, as the vegetation
temperature is the outcome of the energy balance of the surface, the transpiration rate
is related to the balance between radiative forcing and convective and evaporative
cooling.
If the stomata are fully closed (and
r
s
→∞) there may still be some vapour trans-
port through the cuticle. In that case the relevant resistance in Eq. (
6.20
) would be the



























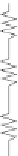


















Search WWH ::

Custom Search