Geoscience Reference
In-Depth Information
CARBONIFEROUS
(1)
(2)
(3)
(4)
(5)
(6)
VI
LN
LE
kockeli
praesulcata
uppermost Famennian
quick changing climate
?
costatus
LL
expansa
VH
?
VCo
styriacus
postera
upper Famennian
climatic
vs
. correlation
challenge
trachytera
velifer
GF
?
G
marginifera
marginifera
rhomboidea
rhomboidea
lower-middle
Famennian
vegetation crisis
C
crepida
crepida
DV
triangularis
triangularis
linguiformis
rhenana
BA
late Frasnian
warm climate
gigas
jamieae
hassi
punctata
transitans
falsiovalis
BM
A. triangularis
BJ
asymmetricus
TCo
GIVETIAN
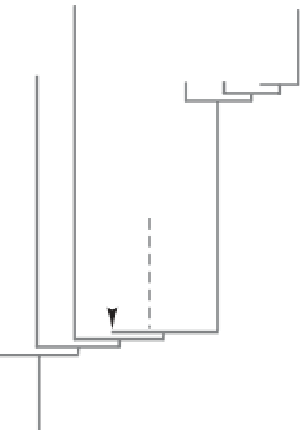
Fig. 2. Chronological, phylogenetic and global (climatic) context of earliest diversification of tetrapods. (1) - (4): as for
Figure 1; (5) phylogenetic relationships (after Ahlberg et al. 2008; Tetrapoda n. gen. et sp. is MGUH VP 6088; taxa
which are not included in this phylogenetic analysis are kept aside on the right; and (6) southeastern Euramerican
climatic conditions after Streel (2007a, b). Note that the so-called 'upper Famennian climatic versus correlation
challenge' is not yet solved due to contradiction in the relation conodont/miospore in the western part (now USA) of the
south Euramerican area.
that tetrapod origins could date back to the earliest
differentiation of these major clades sometime in
the Silurian - Early Devonian. Young considers
this hypothesis as a plausible alternative hypothesis
to the present paradigm of the elpistostegids includ-
ing the sister-group of tetrapods. This alternative is,
of course, strongly supported by the age of the Gle-
nisla trackway. However, the Glenisla trackway
may instead be that of a large arthropod (a type of
misinterpretation already encountered for other tet-
rapod
on an out-of-Euramerica scenario for the origin of
tetrapods (Clack 2002; Blieck et al. 2007).
In seeking an alternative to the consensus
hypothesis, Young (2006) recalled that Late
Devonian tetrapods are very frequently collected
among fish assemblages that include placoderms
(phyllolepids and antiarchs - Remigolepis and
Bothriolepis; Leriche 1931; Clack 2006, table 1
where the locality of Strud, Belgium, has no
placoderm mentioned but did yield placoderms:
Phyllolepis, Bothriolepis, and Groenlandaspis),
acanthodians (gyracanthids a.o.), dipnoans
(Soederberghia being significantly known from
Euramerica and Australia, Ahlberg et al. 2001),
porolepiforms (Holoptychius being the most wide-
spread), osteolepiforms and others. Considering
various hypotheses for the palaeogeographical
origins of those taxa, Young (2006) concludes that
'The various possibilities for faunal connection
(“migration”, “dispersal”) between known tetrapod
localities [his fig. 4] cannot be resolved on the
available evidence' and that we should focus again
'on the possibility of a Gondwanan or Asian origin
for the first land animals'.
tracks,
Clack
1997),
disproving
Young's
hypothesis.
As shown on the phylogenetic relationships in
Figure 2, the most basal tetrapods are from the
Old Red Sandstone Continent (Euramerica). Sinos-
tega, from the Frasnian of China, is too incomplete
to be included in a phylogenetic analysis and Metax-
ygnathus, from the Frasnian or Famennian of Aus-
tralia, is assumed to be more derived than
Densignathus and Ventastega (Ahlberg et al.
2008). Because the sister-group of tetrapods,
namely Panderichthys (Tiktaalik, Elpistostege)or
elpistostegids, is exclusive to the same palaeoconti-
nent (e.g. Daeschler et al. 2006), a consensus arose