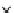Geoscience Reference
In-Depth Information
Area (%)
0 10 20 30 40 50 60 70 80 90 100
0
10
D
TA
20
Area A
Fig. 4.9
An illustration of how the form
factor (
=
the volume development),
Vd
, can be used to express the form,
here given by the relative hypsographic
curve (
=
the depth-area curve) of lakes.
Shallow lakes with a small
Vd
have a
relatively large area above the wave
base (the surface-water areas), where
processes of wind/wave-induced
resuspension will influence the bottom
dynamic conditions. Deep, U-formed
lakes generally have smaller areas above
the wave base (ET areas) and large
deep-water areas. The equation describes
how the accumulation areas (Area A)
may be calculated from data on lake area
(Area), maximum depth (
D
max
), the
depth of the wave base (the critical
depth,
D
TA
) and the form factor (
Vd
).
(Modified from Håkanson 1999.)
30
40
A
=
f
(
Vd
,
D
max
,
D
TA
, Area)
50
60
70
Form
factor (
Vd
)
80
0.6
1.2
1.8
2.4
3
.
D
m
/
D
max
Vd
=
90
100
Area A = Area
.
((
D
max
-
D
TA
)/(
D
max
+
D
TA
.
exp(3 -
Vd
1.5
)))
0.5/
Vd
oxygen status of the sediments. Zoobenthos will
not generally survive if the oxygen concentration
at the sediment-water interface becomes lower
than about 2 mg L
−1
. When the zoobenthos die,
the biological mixing (bioturbation; see Fig. 4.10)
of the sediments will stop. This has profound
consequences for the lamination of the sediments
and, hence, has implications for interpretations
about the age distribution of sediments.
Generally, various types of zoobenthos live in
sediments down to about 5-15 cm sediment depth
(see Fig. 4.11). This upper part of the sediment
column is biologically active in the sense that
the bottom fauna can influence the physical,
chemical and biological conditions of the sedi-
ments and cause bioturbation. Important groups
of bioturbators in lakes are large zoobenthos
such as worms and crustaceans. Benthic animals
are known to display great areal, temporal, ver-
tical and species-specific patchiness within lakes
and great variability among lakes. It has been
shown that bottom animals can eat sediment
up to an average of seven times and this will,
evidently, strongly influence both the age and
age distribution of the sediments (see Håkanson
& Jansson 1983).
If the larger animals (macro- and meiofauna)
die, bioturbation is halted and laminated (layered
and unmixed) sediments may appear. Continu-
ous sedimentation will cause the sediment layer
to grow upward so that the bioturbation limit,
i.e. the limit between the upper biological layer
and the lower biopassive (or geological) layer,
moves upward. As stressed previously, bioturba-
tion will influence the age and the age distribu-
tion at any given sediment depth and this has
evident and profound implications for sediment
dating, i.e. when the sediments are used as a his-
torical archive. When bioturbation is negligible,
for example during anaerobic conditions, the
age of the laminated sediments can be deter-
mined in a very straightforward manner by
'counting varves'. Section 4.4 will discuss some
methods to determine the age of lake sediments
and exemplify why this is important in lake
studies.