Agriculture Reference
In-Depth Information
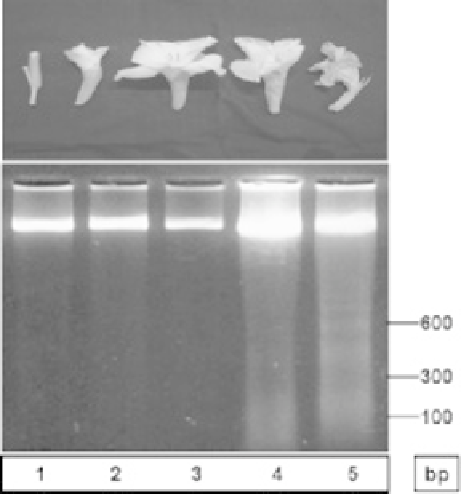
DNA fragmentation detected in petals from various developmental stages in gladiolus (Arora and Singh,
Fig. 5.1
2006).
et al., 2004; Langston et al., 2005), and proteases (Stephenson and Rubinstein, 1998;
Wagstaff et al., 2002; Arora and Singh, 2004) are upregulated during petal senescence
in several species. In
Sandersonia aurantiaca
, a member of a class of cysteine pro-
teases carrying the KDEL C-terminal motif is expressed during senescence (Eason et al.,
2002), which shows homology to a protease from
Ricinus communis
implicated in
ricinosome-mediated endosperm PCD (Gietl et al., 1997). This suggests a mechanism for
petal cell PCD, at least in this species, which may parallel the vacuole-driven autophagous
model, previously described in the
Ricinus
endosperm.
5.5 Senescence signals: Cell sensitivity
The two main correlative events in plant senescence (i.e., pollination versus petal senescence
and grain filling versus leaf senescence) indicate that at least in those cases signals that
initiate the senescence program are produced. Strictly speaking, however, these signals
just hasten or coordinate the senescence program rather than being the real event: petals
eventually senesce even in the absence of pollination (O'Neill and Nadeau, 1997), and
leaves eventually die even without flower or seed forming (Wilson et al., 1992).
It has been proposed that leaf senescence is triggered by age-related decline in photo-
synthetic processes (Hensel et al., 1993). A possible metabolic control of senescence has
also been proposed (Quirino et al., 2000). The onset of senescence in the leaf has been
assigned to a very early point of time when the first reduction in photochemical efficiency
is detected and
cab
transcript levels begin to decline, but no visible sign of senescence and
no expression of
SA
G 12 are observed (Hinderhofer and Zentgraf, 2001). As well, a careful








Search WWH ::

Custom Search