Biomedical Engineering Reference
In-Depth Information
0.50
1
1.5
2
10
0
10
0
10
2
10
2
10
4
10
4
10
6
10
6
10
8
10
8
0.50
1
1.5
2
Fig. 5.24
Plot of theoretical values of gA channel lifetime
τ
th
(arbitrary units, using
τ
th
∼
e
−
(λ
+
−
λ)
F
dis
l
(arbitrary units). The
lower curve
represents
τ
th
using the screened Coulomb theory while the upper
curve represents
τ
th
derived from the elastic bilayer theory, both explained earlier. In both plots
λ
+
−
λ
is considered constant. Both curves are straight lines (in log-lin plot) meaning
τ
th
drops
exponentially with increasing
d
0
,Eq.
5.13
) as a function of the bilayer thickness gA channel length mismatch
d
0
−
k
B
T
l
at the low mismatch level but
τ
th
(from the screened Coulomb
formula) drops even faster (lower curve) as
d
0
−
l
increases due to the inclusion of anharmonic
terms (explained earlier and in Fig.
5.25
) in the energy at high values of mismatch
−
screened Coulomb model calculation hints at the presence of extra damping, due to
higher order anharmonic terms in the energy expression. This better explains why at
high mismatch values, the channel experiences not just destabilization but also struc-
tural transitions (see Sect.
5.4
) due to the energetic cost of the super-heavy bilayer
deformation energy (see Fig.
5.23
). We therefore conclude that although the elastic
bilayer model [
38
,
40
] which yields the deformation energy dependence according
to
2
[
6
,
13
] may be applicable in the small deformation limit, it requires a
modification for values outside this limit. For the same reason, the theory based on
a linear spring approximation for the coupling between the bilayer and gA channels
[
57
], which explicitly shows an exponential damping of gA channel lifetime with an
increasing
d
0
−
∼
(
d
0
−
l
)
l
, can be a very good approximation when
d
0
−
l
is relatively small.
However, when
d
0
−
l
is large enough and the interaction between a gA channel
and the bilayer extends to other next-neighbor lipids in the deformed regions of the
bilayer near the channel, an extension of the elastic model is warranted.
Theoretical and experimental results also show identical trends in the regulation of
the gA channel stability induced by the lipid curvature properties. The energy barriers
modestly change due to the change of the lipid-lipid separation
r
LL
(determined from
the square root of the average lipid cross-sectional area in a bilayer) which changes
as the lipid curvature changes. For simplicity, in the theoretical analysis we have
assumed the same value of
r
LL
for both the lipid-lipid separation and the lipid-
peptide separation at the channel bilayer interface. The energy barrier also shows a
modest change due to the change of lipid charge. To some extent, lipid charge also




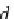





















Search WWH ::

Custom Search