Biology Reference
In-Depth Information
A
B
C
D
MT plus-end tracking
Speckle microscopy
Micromanipulation
Laser ablation
MT growth rate & polarity
Transport & stability
Mechanical properties
MT length & organization
T1
T1
Slow
T1
laser
1
+
-
+
+
-
2
3
4
T2
T2
Fast
T2
1
+
-
+
+
-
3
2
4
MT depolymerization
MT polymerization
Labeled tubulin
Labeled end-
binding protein
Microneedle
Figure 3.3 Methods for measuring MT dynamics and organization within the spindle.
(A) MT end-binding proteins can be fluorescently labeled and used to track the growing
ends of MTs in the spindle. These do not track with the depolymerizing ends of a MT. (B)
MT flux can be measured when fluorescently labeled tubulin “speckles” are incorpo-
rated into a MT. Speckle movement can then be tracked or lifetime within a MT can
be assessed. (C) Micromechanical properties of the spindle can be measured using
force-calibrated microneedles to infer mechanistic and architectural features of the
spindle. (D) Femtosecond laser ablation cuts subsets of MTs in the spindle, leading
to depolymerization from the new plus-end. Organization and architecture of the spin-
dle can be inferred from the depolymerization dynamics.
In
S. cerevisiae
, spindle MTs were observed to turn over from their
plus-ends within the spindle body, but flux through the spindle as in most
metazoan spindles was not observed (
Maddox et al., 2000
). This suggests that
dynamic MTs are a property of all spindles, while poleward MT flux is not.
Furthermore, experimental conditions that block poleward MT flux in cul-
tured cells by inhibiting two MT-depolymerizing proteins did not prevent
spindle assembly or chromosome alignment (
Ganem et al., 2005
). However,
chromosome segregation errors occurred under these conditions, indicating
that MT flux may play a role in promoting robust spindle function, perhaps
by promoting correct k-fiber-chromosome connections. Another more
speculative possibility is that poleward flux is critical, particularly in large








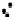











































































































Search WWH ::

Custom Search