Biology Reference
In-Depth Information
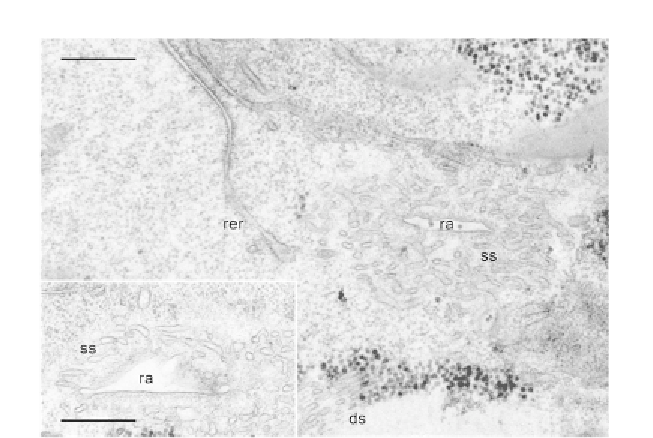
Figure 9.4 Silencing of CRC-II/InsP
3
RinP. tetraurelia causes degradation of the CVC.
Note the unusually loose packing of the smooth spongiome (ss) around a radial arm
(ra). Remnants of the decorated spongiome (ds) are recognized only at a distance from
the smooth spongiome. As in a normal cell (
Fig. 9.2
), some cisternae of the rough ER
(rer), partially devoid of ribosomes, approach the CVC. Inset: Further degradation of
the smooth spongiome around a radial arm which, on its lower part, is kept in shape
by a microtubule support. Again cisternae of the rough ER approach the CVC.
Bars¼0.1 mm. Unpublished micrographs
from experimental
series described by
Ladenburger et al. (2006)
.
indicates mutual dependency of the delivery of CVC-resident proteins—an
interplay between fusion capacity, Ca
2
þ
regulation, and H
þ
sequestration as
the basic primary function of the CVC.
7.4. Hypothetic considerations about de novo CVC biogenesis
During
de novo
biogenesis in
ciliates
, the CVC is placed at predictable sites,
and several scenarios can be discussed. Since both of the new CVCs always
assemble at defined sites of the cell surface and in strict relationship to the
inherited organelles, this speaks for an epigenetic control by morphogenetic
factors. Its formation/expression may depend on the context of the defined
geometrical arrangement of the cortex at a certain distance in anterior direc-
tion from the old CVCs. In analogy to multicellular systems, control may
follow the Gierer-Meinhardt model (
Gierer and Meinhardt, 1972;
Meinhardt, 2006
)—hypothesis (i)—involving unknown soluble factors;
these may operate by an antagonism of stimulatory and/or inhibitory effects.
Search WWH ::

Custom Search