Biology Reference
In-Depth Information
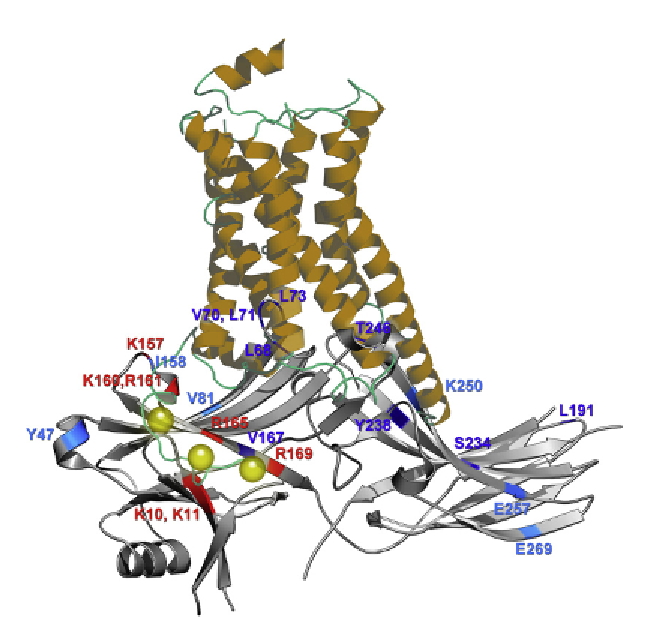
Figure 3.2 Comparison of the cytoplasmic side of active receptor and arrestin. Putative
complex assembled from crystal structure of active
b
2
-adrenergic receptor (from the
complex with G
s
heterotrimer
44
) and arrestin-2.
19
Phosphate-binding residues and
other elements that likely come into direct contact with receptor are shown in red
and blue, respectively. Darker blue shows residues in positions where the mobility of
the engineered spin label is dramatically decreased upon receptor binding, whereas
lighter blue denotes smaller decreases in spin label mobility (based on Ref.
71
). Residue
numbers correspond to bovine arrestin-2 used in.
71
The comparison of these structures
suggests that the receptor-binding surface of inactive arrestin-2 is greater than the cyto-
plasmic part of the receptor resolved in crystal. The receptor C-terminus (not resolved in
any crystal structure) with attached phosphates (yellow spheres) was addedmanually to
position the phosphates near known phosphate-binding positive charges in arrestin.
The analysis of receptor-binding-induced conformational changes in arrestin-1
72
rev-
ealed very small shifts in relative positions of the two arrestin domains, moderate move-
ment of the
“
finger loop
”
toward the receptor, large movement of the neighboring
“
139
loop
”
toward the N-domain and to the side (out of the way of incoming receptor), as
well as the movement of two loops at the distal tips of the N- and C-domain toward
the receptor. Collectively, these rearrangements would allow the finger loop to insert
itself deep into the cavity between receptor helices that opens upon activation, and
move the tips of the arrestin domains closer to the receptor. However, all contacts
expected based on EPR studies of binding-induced changes of spin label mobility in
arrestin-1
73
and -2
71
can only be readily achieved if the receptor helices move even fur-
ther apart than they do in complex with G
s.
44
Search WWH ::

Custom Search