Geology Reference
In-Depth Information
Plate 62 Jurassic Dasyclad Green Algae
Biozonations developed for Tethyan Jurassic and Cretaceous shallow-marine carbonates are based predomi-
nantly on dasyclads and benthic foraminifera.
These zonations use combinations of various stratigraphical concepts. Some zones correspond to taxon range
and concurrent range zones, but many 'coenozones' are assemblage or abundance zones characterized by the
association of several taxa or by a maximum abundance of specific taxa. Coenozone boundaries, therefore, are
not necessarily synchronous time lines.
The Early and Middle Jurassic represent periods of decline and renewal of dasyclads. Common Early Jurassic
and Middle Jurassic dasyclads are
Palaeodasycladus
(-> 1) and
Selliporella
(-> 4). The Late Jurassic, particu-
larly the Kimmeridgian and Tithonian, was characterized by highly diverse and widely distributed dasyclad
associations. Common dasyclads in Late Jurassic-Early Cretaceous times were
Salpingoporella
(-> 2; Pl. 63/7),
Clypeina
(-> 5, 6),
Cylindroporella
(Pl. 63/8) and
Triploporella
. Dasyclads occur in rock-building abundance,
as exemplified by
Clypeina
limestones (-> 5, 6) or limestones with
Campbelliella
(-> 2, 3) in lagoonal or
protected shelf environments.
1
Palaeodasycladus mediterraneus
Pia. Oblique (OS) and cross sections. The alga was common in Tethyan Early and
Middle Liassic (Sinemurian to Domerian) intertidal lagoonal and in outer shelf settings (Sokac 2001) and is used in the
biozonation of Liassic platform carbonates. It characterizes a Late Sinemurian acme zone. Elongated club-shaped thalli,
up to third-order branches diverge with angles of about 45° from the main axis. The sediment consists of poorly sorted
aggregate grains, peloids and micritized algae, sometimes forming nuclei for micrite oncoids (arrows). Note the sparry
patches within the oncoids, indicating the contribution of microbial networks to the formation of 'fenestral oncoids'.
Early Jurassic (Pantokrator limestone, Sinemurian): Korfu Island, Greece.
2
Salpingoporella annulata
Carozzi
.
Oblique (OS) and cross sections (CS). Cylindrical annulated thallus. Pores vertically
flattened, arranged in whorls (arrow). CA: Transversal-oblique section of
Campbelliella
. Open-marine isolated platform.
Late Jurassic (Tithonian): Sulzfluh Range, Graubünden,
Switzerland.
3
Campbelliella striata
(Carozzi)
.
Arrows point to different
sections. These microfossils were formerly regarded as
incertae sedis, pteropod gastropods, large aberrant tintin-
nids, or pallets of boring teredinid bivalves. They are now
recognized as dasyclad algae. The calcareous skeleton con-
sists of a succession of articles interpenetrated into each
other. Each funnel-shaped article (A) contains the pores
of a single whorl (De Castro 1993).
Campbelliella
occurs
in abundance with small benthic foraminifera and
Favreina
in restricted lagoonal platform carbonates. Late Jurassic
(Tithonian): Sulzfluh Range, Graubünden, Switzerland.
4
Selliporella donzellii
Sartoni and
Crescenti. Oblique lon-
gitudinal section. Note double whorls (arrows) with pores
thickening outward (phloiophore). Range of the species:
Bajocian to Late Bathonian. Late Middle Jurassic: South-
ern Apennines, Italy.
5
Clypeina jurassica
Favre. Cross sections. Fertile whorls
shaped like a wide basket and composed of broadening
sporangial chambers. Note (dark) pores in the center of
these chambers. Species range: Late Jurassic to Berria-
sian, common in Kimmeridgian. Late Jurassic: Vrhnica,
Slovenia.
6
Clypeina jurassica
Favre. Isolated whorl fragments. Late
Jurassic: Sulzfluh Range, Graubünden, Switzerland.
-> 1: Flügel 1983
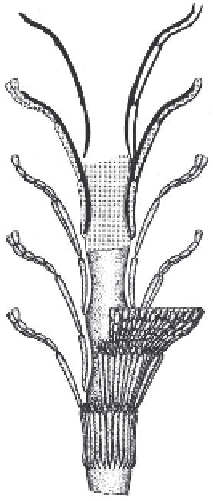
A B
Fig. 10.18.
Reconstructions of Late Jurassic dasyclads.
A
:
Clypeina jurassica
Favre. After Pia (1920).
B
:
Campbelliella striata
(Carozzi). After De Castro (1993).