Geology Reference
In-Depth Information
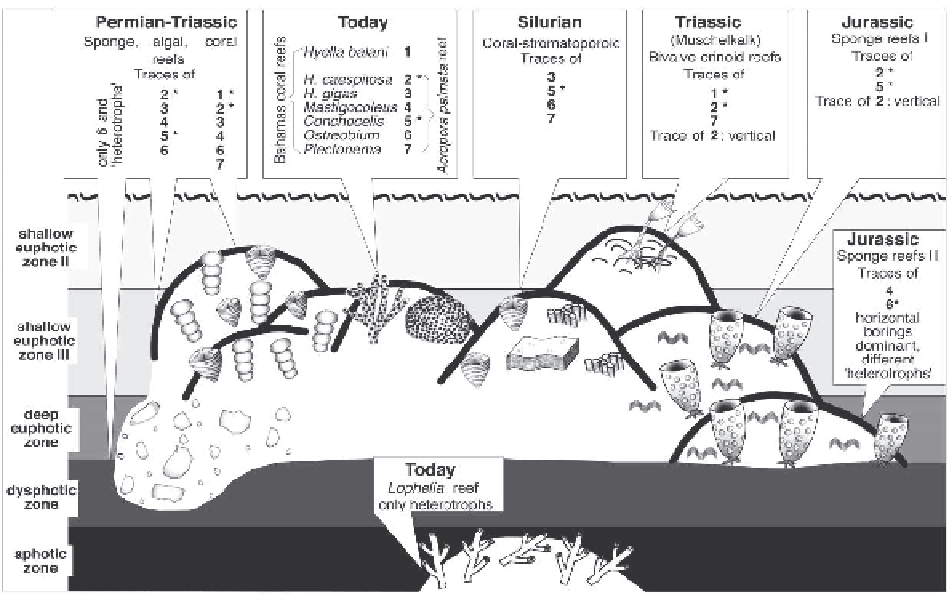
Fig. 9.16.
Bathymetry of different Phanerozoic reef types
inferred from microborer associations. The figure summarizes
investigations of modern shallow-water coral reefs of the Bahamas and deep-water coral reefs of the North Atlantic, and case
studies of ancient reefs from the Silurian of Gotland Island, the Permian of Tunisia, the Middle Triassic Muschelkalk of
Germany and Late Triassic reefs in the Northern Calcareous Alps and Turkey, and Late Jurassic reefs in southwestern Ger-
many.
The bathymetric zones are characterized by (1) the appearance of borings assigned to cyanobacteria (
Hyella, Mastigocoleus
)
versus those affiliated with green algae (
Ostreobium
) or red algae (
Conchocelis
), (2) the frequency of algal-related borings
perpendicular to the surface of the substrate versus those oriented parallel to the substrate surface, and (3) the occurrence of
borings attributed to obligate photoautotrophs (cyanobacteria, algae) versus those produced by obligate or facultative
chemoheterotrophs (fungi).
Each ichnocoenosis is named after key taxa. Not shown in the figure is the shallow eutrophic zone I corresponding to the
supratidal zone. The shallow eutrophic zone II covers the intertidal zone and probably the uppermost part of the subtidal
zone. The shallow eutrophic zone III corresponds to the well illuminated part of the subtidal zone (light intensity 10% of the
surface light). The deep eutrophic zone comprises the less to poorly illuminated subtidal zone (light intensity to about 1% of
the surface light). The dysphotic zone (light intensity to 0.01 to 0.001% of the surface light) is not yet clearly defined by
microborers. Similarly, the microborer associations of the aphotic zone characterized by a dominance of bacteria, fungi as
well as sponge and bryozoan borings need further investigation. The asterisks mark the presence of key taxa of the respective
photic zone.
The use of diagrams showing light intensity versus water depths may lead to a rough estimation of absolute water depths.
But note that light intensity depends on many factors, such as latitude, turbidity of the water, or overshadowing of the bored
substrate by densely growing organisms. Modified from Vogel et al. (1999).
pods became common in Early Cretaceous reefs. Late
Cretaceous rudist reefs exhibit macroborer associations
consisting of bivalves, worms, sponges and cirripedians
(Johnson et al. 2002). Paleogene macroborer associa-
tions are similar to those of the Late Cretaceous. A sig-
nificant change in the abundance and composition of
bioerosion by macroborers took place during the Oli-
gocene and continued in the Miocene. Late Tertiary
reefs exhibit boring patterns similar to recent ones
(Pleydell and Jones 1988; Perrin 2002).
9.3.3.2 Quantitative Changes in the Intensity of
Macroboring in Coral Reefs
Boring intensity can be measured by the semi-quanti-
tative assessment of ichnospecies abundance using field
and laboratory samples, X-ray techniques, serial thin
sections and point-counting (Modern coral reefs: Klein
et al. 1991; Risk et al. 1995; Perry 1998). Preliminary
data provided by Perry and Bertling (2000) suggest that
ancient reefs were variously affected by macroborers