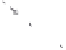Geoscience Reference
In-Depth Information
(a)
0.0
0.1
0.2
BV3, site B, Tussock
BB3, site B, Bare
AH3, site A, Herbs
0.3
(b)
0.1
0.1
0.5
0.5
Caliche layer
Caliche layer
1.00 (m)
1.00 (m)
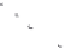
Figure 11.7
The use of dye-tracing studies to demonstrate the effects of plant roots on subsurface flows in storm events:
(a) comparison of tussock grass (
Stipa tenecissima
), bare and small shrubs in Alacant and Murcia, Spain (Cerda, 1997) and (b)
comparison of creosotebush (
Larrea tridentata
) and honey mesquite (
Prosopis glandulosa
) in New Mexico (Martinez-Meza and
Whitford, 1996).
as they grow and extend, and form macropores as the
plant dies (Dunne, Zhang and Aubry, 1991; Devitt and
Smith, 2002; Leffler
et al.
, 2005). Root decay seems to be
relatively rapid in drylands where breakdown is aided
by termite activity (Whitford, Stinnett and Anderson,
1988), and such breakdown typically produces organic
matter, which increases aggregate stability. Dye-tracing
experiments have been used in a number of dry-
land environments to demonstrate the close linkage be-
tween subsurface flow pathways and plant roots (Cerda,
1997; Martinez-Meza and Whitford, 1996; Wang
et al.
,
2007; Ryel
et al.
, 2003, 2004) (Figure 11.7). Archer,
in root structure between shrubs and grasses in Spain pro-
duced a higher apparent hydraulic conductivity in soils
beneath the former.
Because saturation is rarely reached or maintained in
drylands, lateral throughflow is typically rare. Similarly,
vertical percolation is low and in many cases leads to the
development of petrocalcic or other subsurface horizons
(Chapter 7). The difficulty plant roots have in penetrat-
ing these horizons - most likely because of feedbacks
with soil-water chemistry - is often a significant feed-
back on the ability of desert plants to maintain them-
selves (Cunningham and Burk, 1973; McAuliffe, 1994;