Biology Reference
In-Depth Information
A Atlantic halibut
(
Hipoglossus hipoglossus
)
B
Japanese flounder
(
Paralichthys olivaceus
)
TR
a
B
TR
a
A
TR
a
A
TR
b
TR
b
TR
a
B
C
Turbot
(
Scopthalmus maximus
)
D Conger eel
(
Conger myriaster
)
TR
b
1
TR
b
2
TR
a
TR
b
TR
a
A
cTR
a
B
E
Coho salmon
(
Oncorhynchus kisutch
)
F
Pacific bluefin tuna
(
Thunnus orentalis
)
TR
a
bTR
a
A
bTR
a
B
TR
b
bTR
b
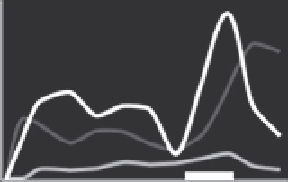
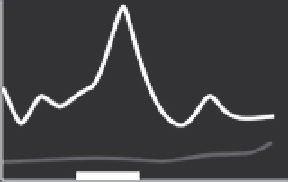
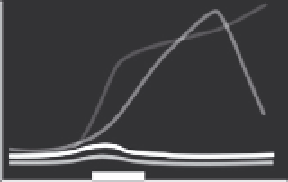
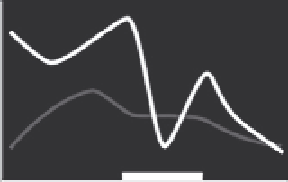
Figure 5.8 Relative different thyroid hormone receptor genes during larval and juvenile
development. Expression is from whole-body assays unless otherwise noted; lines red-
rawn from original figures. The x-axis shows developmental progression and the thick
black bar shows the approximate climax of the metamorphic period. Relative gene ex-
pression in three flatfish: (A) Halibut (
Galay-Burgos et al., 2008
), (B) Flounder (gene ex-
pression in head;
Yamano & Miwa, 1998
), and (C) Turbot (
Marchand et al., 2004
), and
three other teleosts: (D) Eel (
Kawakami et al., 2003
), (E) Salmon (gene expression in brain;
Harada et al., 2008
), and (F) Tuna (
Kawakami, Nozaki, et al., 2008
). The absence of a par-
ticular receptor subtype on a graph does not necessarily mean that the species does not
possess that isoform.
GH and IGF-I are observed during salmonid parr-to-smolt metamorphosis
(
Table 5.2
). Further, treating parr with exogenous GH induces metamorphic
changes, including changes in body shape, pigmentation, and seawater toler-
ance (
Boeuf, 1993; Donaldson, Fagerlund, Higgs, & McBride, 1979; Dufour
& Rousseau, 2007
). Genetic loss of GH likewise delays metamorphic stage
transitions in zebrafish; though this may be a secondary effect of delayed