Biomedical Engineering Reference
In-Depth Information
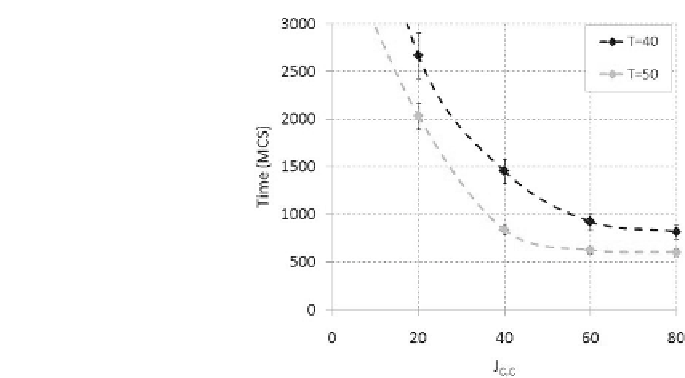
FIGURE 2.8: Time for wound healing as function of the cell{cell adhesion
energy, for T = 40 (dark line) and T = 50 (light line). The error bars show
standard deviations over ten simulations.
in a net energy difference at each copy attempt equal to (1.10). In this case
c(x;t) is the local concentration of HGF, which is assumed to be present ev-
erywhere in the layer under the cells, while the positive parameter
che
C
is the
chemotactic sensitivity, constant in time and homogeneous for all individuals.
Indeed, the chemotaxis coecient is set equal to 0 at cell{cell interfaces, ensur-
ing that chemotactic extensions occur only at cell-ECM surfaces, mimicking
contact inhibition chemotaxis.
The persistent, shape-dependent motion of MLP-29 is modeled with a
further energy term, which coherently is a running mean over the cell past
movements [20, 349]:
H
persistence
=
per
C
jv(t) v(t t)j
2
;
(2.4)
where v(t), defined in Equation (1.12), is an average velocity of the cell, as
t = 60 MCS.
pers
controls the persistence: if
pers
C
= 0 the cells undergoes
uncorrelated Brownian motion, while if
pers
C
is very large their motion is
almost ballistic.
With respect to the model of ARO colonies, the HGF is now explicitly
represented as a continuous object (i.e., a field), whose addition, diffusion, and
absorption are described from a macroscopic point of view, with a standard
diffusion-reaction equation of type (1.1):
@c
@t
= D
n
r
2
c
| {z }
diffusion
"
c
c((
(
x
)
);C)
|
+
{z
}
uptake
c
c(1 ((
(
x
)
);C))
|
+
c
|{z}
addition
;
(2.5)
{z
}
decay







Search WWH ::

Custom Search