Information Technology Reference
In-Depth Information
CA3 CS
CA1 CS
(Hebb, 1949; Marr, 1969, 1970, 1971; McNaughton &
Morris, 1987; McNaughton & Nadel, 1990). This
model provides a framework for associating functional
properties of memory with the biological properties of
the hippocampus. Under this model, the two basic com-
putational structures in the hippocampus are the feed-
forward pathway from the EC to area CA3 (via DG),
which is important for establishing the encoding of new
memories, and the recurrent connectivity within CA3,
which is primarily important for recalling previously
stored memories.
In light of the position of the hippocampus on top
of the cortical hierarchy (figure 9.6), our overall con-
ception of the role of the hippocampus in memory is
one of
binding
together disparate cortical representa-
tions as belonging together in a particular memory. The
detailed “content” of the memory resides out in the cor-
tex, and the hippocampus is simply the binder, group-
ing together subsets of cortical representations. Thus,
the hippocampus does not store
semantic
information,
which is instead represented by the dense interconnec-
tivity and overlapping distributed representations of the
cortex (though it can bind together pieces of semantic
information as isolated “facts” — e.g., “there is a 30%
chance of rain today”).
Hippocampal memories can be characterized as
episodic
— the memory associated with a particular
episode (Tulving, 1972). For example, hippocampal
memories would encode the episode when you heard
the weather report for today. This tendency to repre-
sent specific episodes in the hippocampus makes sense
if we think that its representations are separated out so
that most every “slice” of time gets stored in its own
separate representation. Of course, this is an extreme
view, and it is likely that hippocampal representations
can combine multiple episodes of experience. We also
do not imagine that the hippocampus has a complete
record of all prior “slices” of experience, because these
representations will fade into oblivion if not reinstated
and reinforced periodically.
Because the hippocampus and cortex are so highly
interdependent, many interesting phenomena can be ex-
plained in terms of their interactions. For example, as-
sociations between cortical representations initially en-
coded in the hippocampus can be gradually incorpo-
Entorhinal Cortex
Subiculum
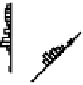
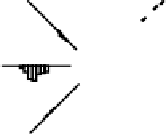
Figure 9.9:
Patterns of firing for neurons in the CA3, CA1,
EC, and subiculum in an eight-arm radial maze, with the
amount of firing shown according to the rat's location and di-
rection of travel within the maze. The cortical-like neurons
in the EC and subiculum have highly distributed activity pat-
terns. In contrast, the CA3 and CA1 neurons are highly sparse
and conjunctive, firing in only one or two specific locations.
ample, figure 9.9 shows the patterns of neural firing
on an eight-arm radial maze, where the neurons in the
CA3 fire only in a particular location on a particular
arm in only one direction. This specificity can be ex-
plained if the CA3 neurons can only be activated by
particular
conjunctions
of sensory features that are only
present in specific locations. This and a wealth of sim-
ilar data suggests that the CA3, and to a lesser extent
the CA1, has sparse conjunctive representations, which
will be very important for our model. In contrast, the in-
put/output areas (EC and subiculum) have a much more
distributed, overlapping character that is typical of the
cortex.
9.3.2
Basic Properties of the Hippocampal Model
Our model is based on what McNaughton has termed
the “Hebb-Marr” model of
hippocampal function


















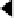



































































Search WWH ::

Custom Search