Information Technology Reference
In-Depth Information
These activation states are local to the synapse where
the weight changes must occur, and we will see below
how such weight changes can happen in a biological
synapse.
There are a number of important properties of the
GeneRec learning rule. First, GeneRec allows an error
signal occurring anywhere in the network to be used to
drive learning everywhere, which enables many differ-
ent sources of error signals to be used. In addition, this
form of learning is compatible with — and, moreover,
requires — the bidirectional connectivity known to ex-
ist throughout the cortex, and which is responsible for a
number of important computational properties such as
constraint satisfaction, pattern completion, and attrac-
tor dynamics (chapter 3). Furthermore, the subtraction
of activations in GeneRec ends up implicitly computing
the derivative of the activation function, which appears
explicitly in the original backpropagation equations (in
equation 5.24 as
h
j
(1h
j
)
). In addition to a small gain
in biological plausibility, this allows us to use any arbi-
trary activation function (e.g., the point neuron function
with kWTA inhibition) without having to explicitly take
its derivative. Thus, the same GeneRec learning rule
works for virtually any activation function.
The next section presents the derivation of equa-
tion 5.32, and the subsequent section introduces a cou-
ple of simple improvements to this equation.
o
k
. . .
∆
w
jk
=
h
j
−
(t
k
−o
k
)
h
j
. . .
h
+
−
h
j
−
∆
w
ij
=
(
)
s
i
−
s
i
. . .
External Input
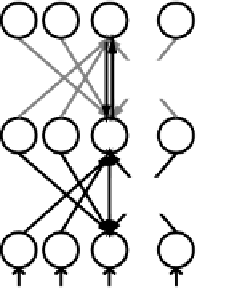
Figure 5.10:
Weight updates computed for the GeneRec al-
gorithm.
phase). In particular, the hidden units need to receive
the top-down activation from both the minus and plus
phase output states to determine their contribution to the
output error, as we will see in a moment.
As usual, we will first discuss the final learning rule
that the GeneRec derivation produces before deriving
the mathematical details. Conveniently, the learning
rule is the same for all units in the network, and is es-
sentially just the delta rule:
(5.32)
for a receiving unit with activation
y
j
and sending
unit with activation
x
i
in the phases as indicated (fig-
ure 5.10). As usual, the rule for adjusting the bias
weights is just the same as for the regular weights, but
with the sending unit activation set to 1:
5.7.1
Derivation of GeneRec
Two important ideas from the recirculation algorithm
allow us to implement backpropagation learning in a
more biologically plausible manner. First, as we dis-
cussed above, recirculation showed that bidirectional
connectivity allows output error to be communicated to
a hidden unit in terms of the difference in its activa-
tion states during the plus and minus activation phases
(5.33)
If you compare equation 5.32 with equation 5.21
from backpropagation (
w
ij
= Æ
j
x
i
), it should be
clear that the phase-based difference in activation states
of the receiving unit
(yj yj
, rather than in terms of the
Æ
's multiplied
by the synaptic weights in the other direction. Bidirec-
tional connectivity thus avoids the problems with back-
propagation of computing error information in terms of
is equivalent to
Æ
in
backpropagation. Thus, the difference between the two
phases of activation states is an indication of the unit's
contribution to the overall error signal. Interestingly,
bidirectional connectivity ends up naturally propagat-
ing both parts of this signal throughout the network, so
that just computing the difference can drive learning.
, sending this information backward from dendrites,
across synapses, and into axons, and multiplying this
information with the strength of that synapse.
Search WWH ::

Custom Search