Information Technology Reference
In-Depth Information
−
Dendrite
+
Θ
Θ
Postsynaptic
complex
chemical
processes
2+
[Ca ]
i
NMDA
2. Glutamate released with
spike binds to and opens
NMDA channels
3. Ca++ enters through
NMDA channels
Figure 4.2:
Relationship between LTP and LTD, where
a moderate amount of increased intracellular calcium
(
[Ca
++
]
i
) leads to LTD, but a larger amount leads to LTP.
1. Mg+ unblocks NMDA
channels as postsynaptic
V_m increases
Ca++
nature of these complex chemical cascades (Lisman,
1989, 1994; Bear & Malenka, 1994). Further, this form
of LTD is consistent both with the LTD necessary for
model learning, and with that necessary for task learn-
ing, as will be explained below in section 5.8.3.
Although the NMDA-mediated LTP/D mechanism is
relatively simple and consistent with a large amount
of data, the biological picture may be somewhat more
complicated, for a number of reasons:
Presynaptic
Axon
Figure 4.1:
Sketch of biological mechanisms that lead to
LTP/D. NMDA channels open when the postsynaptic mem-
brane potential (
V
m
) is sufficiently elevated and glutamate is
being released by the presynaptic neuron. This allows calcium
ions into the postsynaptic neuron, which triggers a cascade of
complex chemical processes that ultimately result in the mod-
ification of synaptic efficacy.
Other receptors, channels, and neurotransmitters may
play a role in LTP/D. For example: (a) there is evi-
dence for the involvement of the metabotropic gluta-
mate (mGlu) receptors in LTP (Bashir, Bortolotto, &
Davies, 1993; Bear & Malenka, 1994), (b) there are
a number of other ways in which calcium can enter
the postsynaptic neuron (e.g., non NMDA voltage-
dependent calcium channels), and (c) LTP/D may be
regulated by a number of other modulatory neuro-
transmitters (dopamine, serotonin, etc.) in ways that
are not understood or appropriately manipulated in
experiments (see section 6.7 for a more detailed dis-
cussion of the role of dopamine in learning).
to which factors are important is far from resolved, but
it does appear that both presynaptic and postsynaptic
factors could be involved in LTP/D. However, our com-
putational learning mechanisms depend more on the na-
ture of the events that trigger this synaptic modification
than on the mechanisms that actually implement it.
LTD is somewhat less well understood. One promi-
nent idea is that LTD occurs when the synapse is active
at a somewhat lower level than that required to trigger
LTP (Artola, Brocher, & Singer, 1990). Thus, one ex-
planation of this finding would be that the less effective
opening of the NMDA channels (e.g., fewer channels
opened or opened for a shorter time) results in a lower
concentration of calcium ions, which triggers a differ-
ent cascade of chemical processes that ultimately end
up
reducing
synaptic efficacy instead of enhancing it.
Figure 4.2 shows an illustration of this relationship be-
tween LTP and LTD, where the threshold for LTD is
given by
, and the higher LTP threshold is
+
.This
relationship is consistent with one hypothesis about the
There is little evidence that the standard 100Hz for
1 second's activity required for LTP induction occurs
with any reliability in the natural activation patterns
of the cortex. More generally, LTP/D may be sensi-
tive to particular combinations of inducing activation
signal properties (timing, intensity, frequency, dura-
tion, etc.) in ways that have not been explored empir-
ically (indeed, this kind of timing sensitivity is impor-
tant for achieving biologically plausible error-driven
learning, as we will see later).






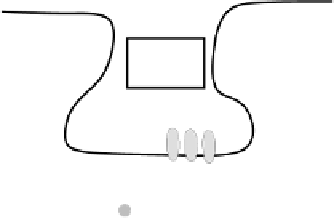



Search WWH ::

Custom Search