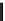Biology Reference
In-Depth Information
Erythrose-4-P
+
phosphoenolpyruvate
Cytoplasm
Plastid
HCT
4-Coumaroyl-shikimate
Caffeoyl-shikimate
HCT
C3
¢
H
Shikimate
Shikimate
Shikimate
4-Coumaroyl-CoA
Caffeoyl-CoA
Chorismate
C4H
Flavonoids
Coniferaldehyde
F5H
H-lignin
P
h
e
Trp
Arogenate
Cell wall
Tyr
Phe
G-lignin
S-lignin
Fig. 3. Connection between the shikimate and phenylpropanoid pathway. Shown
is an outline of the shikimate pathway (pale green box) and the phenylpropanoid
pathway. Only branch-point metabolites are given. Note the dual function of shiki-
mate both as a far upstream precursor of 4-coumaroyl-CoA and as a cofactor in the
HCT/C3
0
H-mediated 3-hydroxylation yielding caffeoyl-CoA.
irreversible step towards S- and G-lignins (
Anterola and Lewis, 2002
). Thus, it
is the 'gatekeeper at a point of no return' to the bulk of lignin in angiosperms,
which is quantitatively by far the biggest sink of carbon channelled through the
shikimate pathway, especially in woody species. But obviously, a sufficient
pool size of aromatic amino acids and general phenylpropanoid pathway
intermediates must be ensured at all times to allow production of quantitative-
ly 'less important' but nevertheless pivotal compounds including proteins and
non-lignin plant natural products. Thus, it is a tempting hypothesis to assume
that using the upstream precursor shikimate as a cofactor is an immediate
metabolic regulatory mechanism: as soon as shikimate pool sizes drop below a
threshold value, the committed step in S- and G-lignin biosynthesis becomes
devoid of its cofactor and thus shuts down. This would allow maintaining
necessary aromatic amino acid pool sizes in the presence of a massive sink
pathway. Such a regulatory mechanism would act almost instantly and would
not require transcriptional regulation or protein modifications. However, it
will be difficult to proof this hypothesis, as any manipulation interfering with
the production of shikimate used as a cofactor will also interfere with the
production of the 'substrate' shikimate. This hypothesis also assumes equilib-
riumbetween shikimate pool sizes in the chloroplast, the locale of the shikimate
pathway, and the cytosol, where the phenylpropanoid pathway is localized.
Presumed shikimate transporters would be obvious targets to distinguish