Biology Reference
In-Depth Information
the '
nif
'-gene cluster intact along with 22 genes governing heterocst differentiation, it has become
an obligatory confi nement for the endosymbiont to fi x nitrogen and release it to the host. It is also
unable to reassimilate the released nitrogen due to the absence of transport mechanisms. So the host
environment becomes very restrictive and the host supplies only limited supply of carbohydrates
and phosphate so that the growth of the symbiont is slowed down (Ran
et al.,
2010). The affected
metabolic processes in the symbiont
N
.
azollae
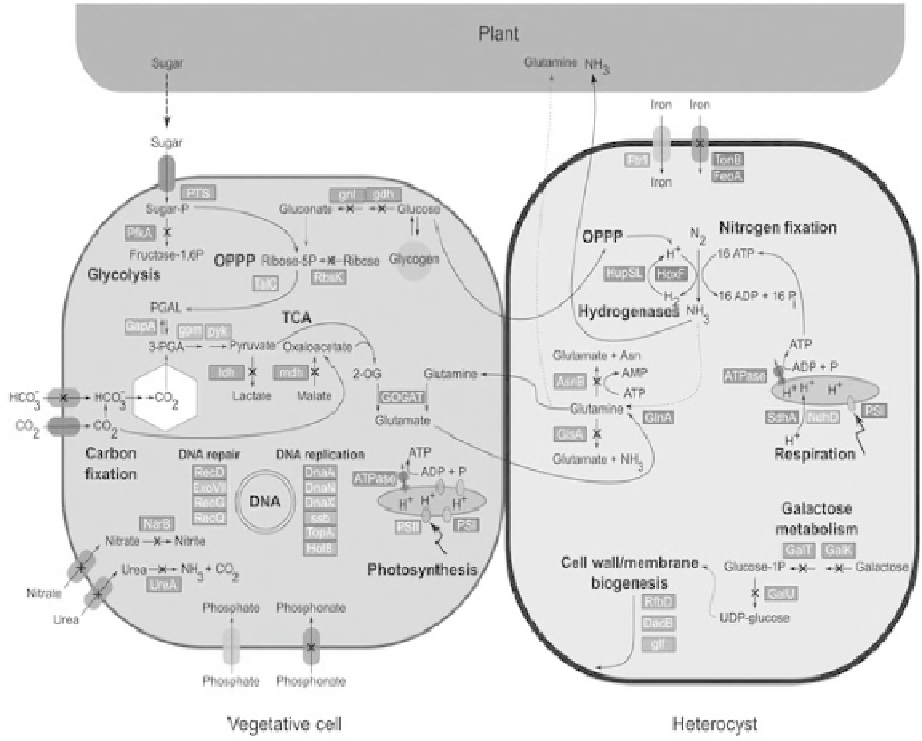
0708 have been depicted in Fig. 10.
Figure 10:
Schematic illustration of important metabolic and genetic information pathways in
Nostoc azollae
. The left cell
represents a vegetative cell while the right a nitrogen-fi xing heterocyst. Red color indicates pseudogenes lacking a functional
counterpart in the
N
.
azollae
genome. Orange indicates pseudogenes where a functional counterpart is present elsewhere
in the genome. Fully functional gene(s) are illustrated (blue) only if their function is linked to other processes in the fi gure.
The localization of pathways in vegetative cells or heterocysts is representative only for nitrogen fi xation (heterocysts) and
PSII activity (vegetative cells). Note that only a minor part of the nitrogen fi xed in heterocysts is incorporated using the
GS-GOGAT pathway and used for synthesis of amino acids, while most is exported to the plant as NH
3
. Sugar is provided
by the plant in an as yet unknown form; putatively imported via the sugar phosphotransferase system. Function has been
lost in the glycolytic pathway as the
pfkA
gene, encoding 6-phosphofructokinase, is a pseudogene and sugar metabolism in
the
Azolla
cyanobiont probably proceeds via the oxidative pentose phosphate pathway. Extensive loss of function is evident
among genes involved in uptake and transport of nutrients and
N
.
azollae
has lost the capacity to both import and metabolise
alternative nitrogen sources. With the kind permission of B. Bergman [Ran
et al
. (2010)
PLoS ONE
5(7):
e11486. doi:10.1371/
journal.pone.0011486] doi:10.1371/journal.pone.0011486.g006.