Biology Reference
In-Depth Information
strain PCC 6803 (Table 4). With a G+C content of 40%, the number of genes in
C
.
raciborskii
CS-505 is
3,968 of which 3,452 constitute the coding sequences (Stucken
et al
., 2010). In case of
R
.
brookii
D9 the
total number of genes is 3,088 of which 3,010 comprise the coding sequences. With the number of genes
for rRNA and tRNA being 9 and 42, respectively in both the organisms, the genome of
C
.
raciborskii
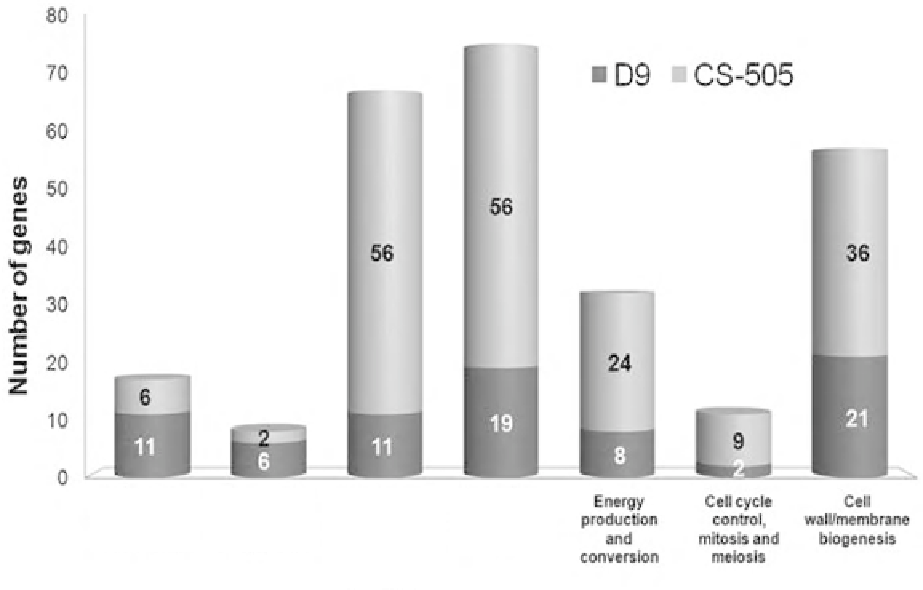
CS-505 additionally possesses large number of genes for transposases, phage integrases and repeated
regions (Fig. 6). A comparison of the sequenced genomes of heterocystous cyanobacteria led to the
identifi cation of COGs for fi lament formation (32 genes), diazotrophic metabolism (49 genes) and
heterocyst development (149 genes) and nitrogen fi xation (49 genes). The difference in the number of
genes required for fi lament formation though appeared to be small in between
C
.
racoborskii
CS-505
(23 genes) and
R
.
brookii
D9 (20 genes), the common set of genes required for fi lament formation has
further been reduced to 10 genes by the inclusion of unfi nished genomes of
Nostoc azollae
078 and
M
.
chthonoplastes
PCC 7420 in the sequence comparison. The number of genes required for diazotrophy
in
C
.
raciborskii
CS-505 is 38 but in
R
.
brookii
D9 only fi ve of these are present. The core number of
orthologues accounted for 10 genes. Likewise, the COGs for heterocyst development have been found
to be 149, among these 58 unique genes have been found that are common to all heterocyst-forming
species and this number is further reduced to 41 if
N
.
azollae
078 is included for sequence comparison.
80
70
D9
CS-505
60
50
40
30
20
10
0
Cell cycle
control,
mitosis and
meiosis
Cell
wall/membrane
biogenesis
Amino acid
transport and
metabolism
Coenzyme
transport and
metabolism
Secondary
metabolites
biosynthesis,
transport and
catabolism
Replication,
recombination
and repair
Energy
production
and
conversion
Figure 6:
Distribution of the unique coding squences (CDS) of
Cylindrospermopsis raciborskii
CS-505 and
Raphidiopsis brookii
D9 into Cluster of Orthologous Groups (COGs). Only COG categories over-represented by CDS of
C. raciborskii
CS-505 or
R.
brookii
D9 are shown. Unique CDS were obtained by a bidirectional best hits (BeTs) search between both genomes using a 30%
cutoff. With the kind permission of M. Vásquez, Department of Molecular Genetic and Microbiology, Faculty of Biological
Sciences, Pontifi cia Universidad Católica de Chile, Santiago, Chile & Millenium Nucleus EMBA, Santiago, Chile, [Stucken
et al
. (2010)
PloS ONE
5(2):
e9235; doi:10.1371/journal.pone.0009235] doi:10.1371/journal.pone.0009235.g002