Biomedical Engineering Reference
In-Depth Information
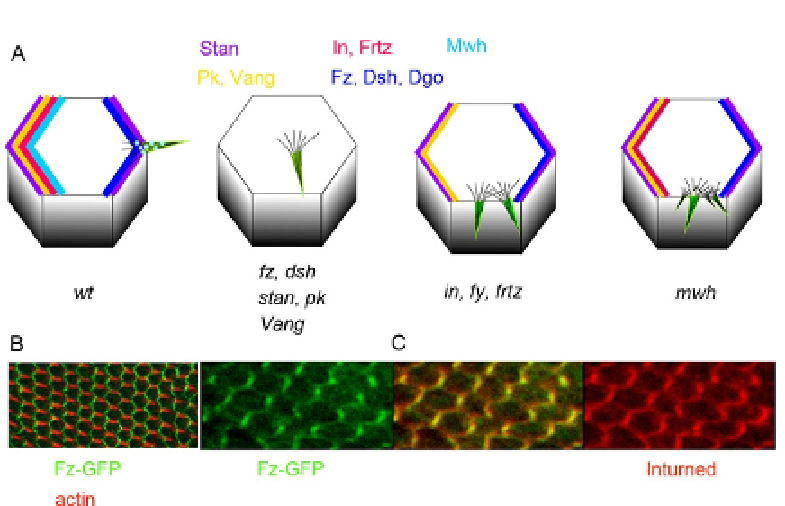
Figure 1.2 Polarized protein accumulation and mutant phenotypes. A shows cartoons
of wild-type and mutant cells. In the wild-type cell, all of the fz/stan pathway proteins
accumulate asymmetrically and hairs are formed at the distal vertex. In cells, mutant for
fz or other core genes hairs form at a relatively central location and no proteins accu-
mulate asymmetrically. In cells, mutant for in or any of the other PPE genes multiple
hairs form at an abnormal location on the cell periphery. The core proteins accumulate
asymmetrically but the PPE proteins and Mwh do not. In cells, mutant for mwh genes
multiple hairs form at an abnormal location on the cell periphery. The core and PPE pro-
teins accumulate asymmetrically but Mwh does not. B shows the accumulation of
Fz-GFP by direct imaging of GFP (green) (no immunostaining) and actin in red (hairs).
C shows the coordinate asymmetric accumulation of Fz (green) and In (red).
8 photoreceptor cells. The arrangement of the photoreceptor cells is asym-
metric and chiral giving each ommatidia and the eye as a whole a polarity
(
Fig. 1.4A
). As each ommatidia develops, there is a stepwise recruitment
of photoreceptor cells (for a recent review of this, see
Kumar, 2012
). The
R3 and R4 cells are recruited at the same time. The cell located closer
to the equator becomes R3 and the other R4 (
Fig. 1.4A
). The ommatidia
undergo a subsequent rotation and this is guided by the position of the R3
and R4 cells (
Fig. 1.4B
). The
fz
pathway controls PCP by regulating the
R3/R4 cell fate decision (
Zheng et al., 1995
). This is based on the R3 cell
having higher Fz activity and R4 higher Vang (
Strutt, Johnson, Cooper, &
Bray, 2002
). This leads to the Delta-dependent activation of the Notch (N)
receptor in R4, which controls cells fate (
Cooper & Bray, 1999; Fanto &
Mlodzik, 1999; Tomlinson & Struhl, 1999
). There is an argument in the