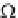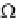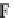Biomedical Engineering Reference
In-Depth Information
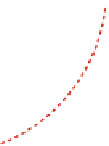
Fig. 14.1
Sketch of the
active strain decomposition
entailing an intermediate
virtual configuration
Ω
e
between the reference state
Ω
o
and the current
configuration
Ω
. Similar
splittings have been proposed
in finite elastoplasticity (Lee
and Liu,
1967
), growth and
material remodeling (Taber
and Perucchio,
2000
; Menzel
and Waffenschmidt,
2009
),
and mechano-chemical
interactions (Murtada et al.,
2010
)
Perucchio (
2000
); Nardinocchi and Teresi (
2007
), assumes that the deformation gra-
dient
F
can be rewritten in terms of a Lee-type multiplicative decomposition (Lee
and Liu,
1967
), i.e.
F
=
F
e
F
o
,
(14.5)
where
F
o
is the active deformation, to be constitutively prescribed in terms of ionic
species concentration, and
F
e
is the passive elastic deformation (see Fig.
14.1
).
Whichever approach that is chosen, the model should satisfy due mathematical
properties (such as frame indifference and ellipticity of the total stress), and the con-
stitutive laws need to recover physiological relevant behaviors (such as the Frank-
Starling effect, where an increase of chamber volume at end-systolic pressure and
stroke work reflects on the tissue as a monotonic increase in isometric tension), (Lee
and Liu,
1967
).
Comparisons between the usual active stress method and the active strain ap-
proach from a numerical viewpoints has been carried out in Rossi et al. (
2012
).
Defining the variables
γ
f
,
γ
s
,
γ
n
as the relative displacements in the directions
f
0
,
s
0
,
n
0
, (fibers, sheets and sheets-normal directions) of a single cell, respectively, the
local
deformation is
F
o
=
I
+
γ
f
f
0
⊗
f
0
+
γ
s
s
0
⊗
s
0
+
γ
n
n
0
⊗
n
0
.
(14.6)
Note that
γ
f
represents the active shortening of the cardiomyocytes, whereas
γ
s
,
γ
n
will take into account the associated thickening, in order to satisfy the incompress-
ibility of the cell itself (Iribe et al.,
2007
;Smerupetal.,
2009
). In Nardinocchi and
Teresi (
2007
), Evangelista et al. (
2011
) and Nobile et al. (
2012
), the contribution
of the terms depending on
γ
s
and
γ
n
are not included. Analogously, for some acti-
vation models (see, e.g., Göktepe and Kuhl,
2010
; Rausch et al.,
2011
), the active
tension is assumed to act exclusively along the fibers direction. However, biaxial
tests provide a measure of the active contributions in the transverse direction. This
quantification can be obtained by either measuring the different rates of calcium