Biology Reference
In-Depth Information
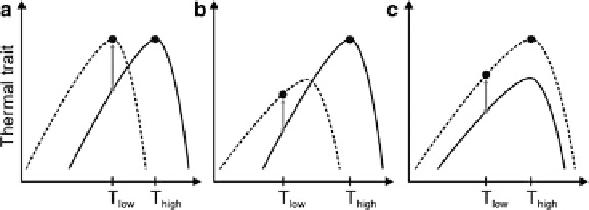
Fig. 3.2
Two individual temperature-response curves of organisms grown at a high temperature
T
high
(
solid line
) and at a low temperature
T
low
(
dashed line
). The
circles
mark the physiological
rate at the respective growth temperature. The
gray arrows
indicate (partial) acclimation at
T
low
.
(
a
) “Complete low-temperature acclimation”, (
b
) “Beneficial acclimation hypothesis”, and
(
c
) “Colder is better hypothesis”
e.g., diurnally or seasonally. The quantitative and/or qualitative metabolic adjust-
ments of proteins that allow temperature compensation are similar to the genotypic
evolutionary adjustments (Clarke
2003
). Also, membranes can undergo changes in
both lipid composition and degree of unsaturation of fatty acids in response to
temperature changes. In this way, they can maintain a relatively constant fluidity
and can alter the state transition temperature (Los and Murata
2004
). These
homeoviscous changes are very important in establishing the appropriate mem-
brane physical state for some enzymes at different temperatures.
Phenotypic acclimation can be tested experimentally by comparing temperature-
response curves (short-term effects) of organisms grown at different temperatures
(long-term effects). The following three parameters of the short-term temperature-
response curves are the most important: maximum rate, temperature optimum,
and performance breadth. Both a shift in the response curve (including temperature
optimum) and increase in maximum rate may be beneficial and maximize perfor-
mance over a broad temperature range. Various acclimation hypotheses are
discussed in evolutionary biology (Deere and Chown
2006
). “Complete tempera-
ture compensation” maintains the physiological rates as the temperature changes
(Fig.
3.2
). However, partial compensation is apparently more common and is
predicted from the “Beneficial acclimation hypothesis” caused by some co-
variation between optimum temperature and maximum rate and some overlap of
the performance curves (Fig.
3.2b
). Temperate populations of
Valonia utricularis
(Chlorophyta, Eggert et al.
2006
) and
Asparagopsis taxiformis
(Rhodophyta,
Padilla-Gamiˇo and Carpenter
2007
) displayed seasonal (partial) temperature
acclimation of photosynthesis, whereas tropical individuals of both species did
not. Alternatively, the hypothesis “Colder is better” predicts that an organism
acclimated to a low-temperature environment will always have a performance
advantage over another organism held at a warmer environment (Fig.
3.2c
). In
this case, the optimal temperature does not change, but the performance curve
overall increases. This type of acclimation of photosynthesis has been shown for the
Antarctic seaweeds
Ballia callitricha
and
Gymnogongrus antarcticus
(Eggert and