Biology Reference
In-Depth Information
(a)
(b)
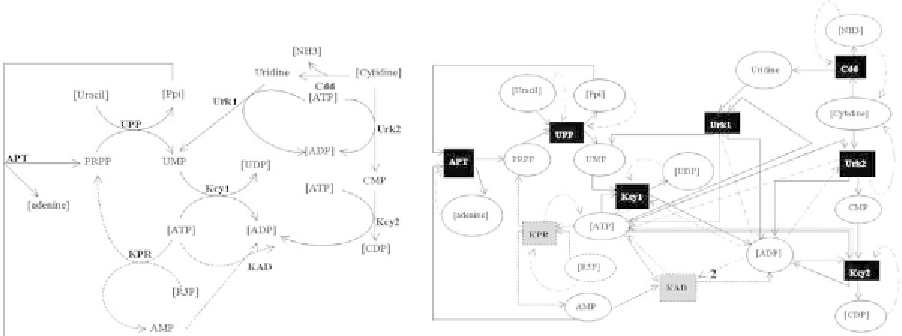
Fig. 7. Part of nucleotide metabolism. The symbols stand for abbreviations of metabolites and enzymes usual in biochemistry.
External metabolites are written in brackets. If the reactions indicated by dashed arrows are absent, the conservation relation ATP
+
ADP
=
const. holds. We do not consider R5P be part of the system in this case. A) Traditional biochemical representation.
B) Petri net representation. The external metabolites were modelled with self-loops, depicted by dotted arrows.
way to not “consume” the input-place tokens and to not “produce” final-place tokens,
M
(
p
)=
M
(
p
)
∀
F
(1)
where I is the set of initial places and F is the set of final/terminal places. Note that the firing rule for the
internal metabolites is
M
(
p
)=
M
(
p
)
−
p
∈
I
∪
F.
(2)
Another possibility is to connect sink places with source places by additional transitions so that a circular
flow occurs [Heiner
et al.
, 2000]. However, it is difficult to find which places have exactly to be
connected with each other, because such transitions could impose unrealistic constraints on the flow
ratios. For example, one cannot regenerate carbon atoms from outgoing nitrogen atoms. A solution can
be to use coloured Petri nets, in which different atom groups can be modelled by tokens of different
colour. Starke [1990] proposed, as another way of description, not to include the initial and final places
in the net. Thus, the boundary is made up of transitions without presets or without postsets. The initial
transitions do not need any tokens to fire. In the traditional modelling of metabolic networks, a similar
description is indeed sometimes used for external metabolites that are of minor importance, such as
inorganic phosphate, water, protons etc. However, applying this technique to all external metabolites has
the drawback that they are not made explicit so that overall molar yields cannot be computed.
Here, we propose an alternative method. For each initial place, we add an arc feeding from the
transition back to this place (Fig. 7) and use the firing rule (2) both for internal and external metabolites.
This guarantees that the number of tokens in the initial places remains unaltered. For each final place,
we add an arc feeding from this place back to the transition producing it. To guarantee that the transition
can always fire, at least one token should be put in the final place at the beginning. However, one should
be aware that this generates self-loops, so that the Petri net is no longer pure. Thus, as far as the external
metabolites are concerned, the incidence matrix does not equal the stoichiometry matrix. This is no
problem since the external metabolites are not usually included in the stoichiometry matrix [Heinrich
and Schuster, 1996].
pre
(
p, t
)+
post
(
t, p
)
∀
p
∈
P
−
I
−