Biology Reference
In-Depth Information
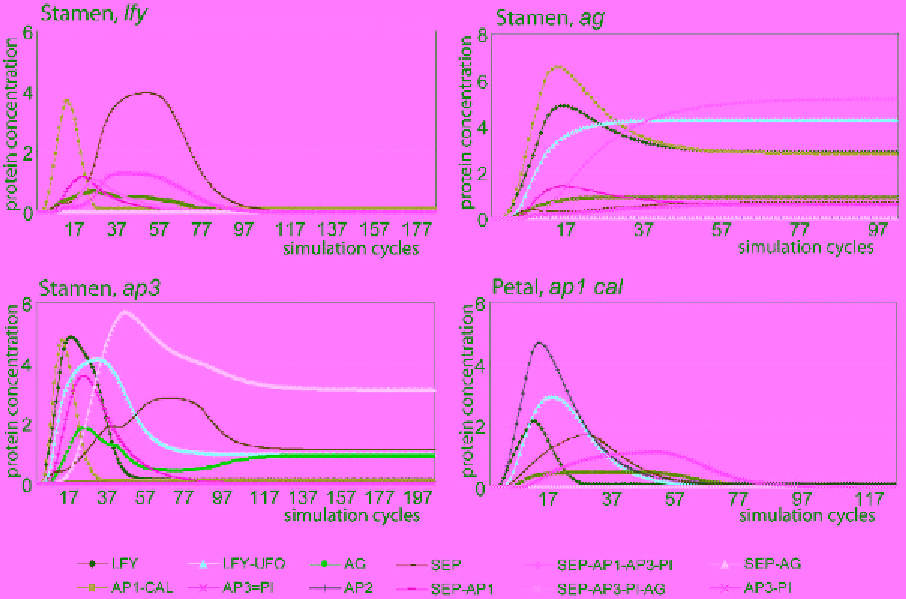
Fig. 5.
Simulation of the network under different mutant contexts.
The mutant described in each plot was simulated by
setting the translation speed of the mutated gene to 0.
The steady state expression profiles coincide with the phenotypes of
experimentally characterized mutants.
dual role of
AP1
and
CAL
as flower meristem identity genes. Thus, the model predicts that different
higher-order complexes may transiently coexist at early time-points of flower meristem development.
In order to further test our model, simulations of various known mutants were performed. The networks
of mutant scenarios were simulated by setting the translation speed of the targeted entity to 0. As can
be seen in Fig. 5, mutation of the
LFY
gene under the stamen conditions results in a transient low-level
activation of AP1/CAL and SEP proteins, which is not sufficient to trigger persistent upregulation of
floral homeotic genes. In agreement with this, the
lfy
mutant produces leaf-like organs instead of stamens.
Similar results were obtained for the other floral organs. In contrast,
ag
and
ap3
mutants form petal
(AP1/SEP/AP3/PI) and carpel (AG/SEP) complexes in the third whorl, respectively, which is consistent
with the homeotic mutant phenotypes that are described in the literature [Bowman
et al.
, 1989; Yanofsky
et al.
, 1990; Jack
et al.
, 1992].
Network robustness and response to stochastic oscillations
The response of the network to stochastic oscillations was measured by setting the value of independent
entities to oscillate at random within a given interval during the simulation time. It was observed that
the requirement of protein complexes as end regulators of the network leads to dampening of noise and
to a more stable concentration of the complexes themselves. As a result, the regulatory effect of these