Geology Reference
In-Depth Information
sets that display low angle dipping foresets and may be
used as evidence of tidal infl uence (Santos and Rossetti
2006
) . Bidirectional tidal fl ow can create distinct
cross-lamination (ripples) or cross-bedding (dunes).
Sets of ebb-oriented cross-laminae, bounded by fl ood-
oriented cross-laminae (or vice versa) are known as
herringbone cross
-
stratifi cation
and are a good indica-
tor of tidal deposition and may be seen in deep subtidal
portions of a channel. Degree of symmetry in the her-
ringbone structures provides insight into tidal asym-
metry at the point of deposition in time and space. If
one tidal current is weaker than the other, the subordi-
nate current may create a 'cap' of smaller oppositely
directed foresets at the crest of the bedform created by
the dominant current (Mowbray and Visser
1984
) .
However, the complex recirculation and fl ow-segrega-
tion, which occur in most braided or meandering chan-
nels, can create sets of exclusively fl ood- or ebb-oriented
cross-stratifi cation in shallower regions of the channel.
In low-energy settings, such as small channels with
slower fl ows (~0.3 m/s), bedforms are unlikely to form
but parallel laminations may be seen where mud settles
out of suspension during low-fl ow periods and sand is
moved as bedload during times of faster fl ow.
The crests of tidal pointbars are often heavily popu-
lated with worms, mollusks and burrowing crusta-
ceans. A high level of bioturbation is a notable feature
of intertidal regions, providing differentiation between
tidal and fl uvial systems, where infauna are scarce.
Species diversity increases inland from saline to brack-
ish environments (Barwis and Hayes
1979
). Using this
information, in hypersynchronous systems, where
similar tidal ranges can exist at two or more sites, ich-
nology can help to differentiate between regimes based
on species tolerance to salinity and diversity.
Bioturbation differs with position in the tidal range;
below mean low water, bioturbation is relatively sparse,
decreasing into the channel thalweg. Likewise, in
regions of recent slumping, bioturbation may be less
frequent. In the upper regions of a tidal pointbar, how-
ever, faunal activity can be intense. Pearson and
Gingras (
2006
) observed burrow densities of up to
60,000 burrows/m
2
in the upper-intertidal zone of a
muddy pointbar in the Bay of Fundy. Ichnological
investigation showed different assemblages across the
bar (in the upper-subtidal and lower-intertidal zones of
Polykladichnus
- and
Skolithos
-like traces character-
ized the pointbars; Arenicolites-, Diplocraterion-,
Polykladichnus-,
Palaeophycus
-, and
Planolites
-like
forms were found in the middle-intertidal portions of
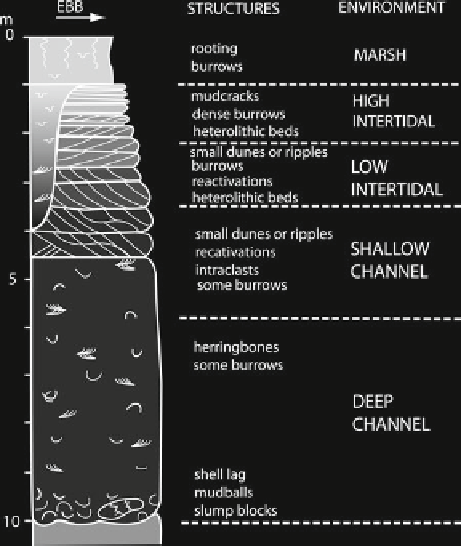
Fig. 11.16
(
a
) A general pointbar facies model (Barwis and
Hayes
1979
)
the pointbars; and in the upper-intertidal, S
iphonichnus
-
and
Polykladichnus
-like burrows were found). These
assemblages are consistent with brackish water
conditions.
Tidal facies are more likely to be preserved when
bioturbation is low. This would be the case in channels
where the thalweg and pointbars have a higher sand
content, as muddy sediment supports more active
infauna. Likewise regions of moderately high veloci-
ties also discourage faunal activity and stratigraphy is
more likely to be preserved (Ashley and Zeff
1988
) . In
regions with low deposition rates, the activity of bur-
rowers may completely obscure bedding (Barwis and
Hayes
1979
; Pearson and Gingras
2006
) . However, if
rates of deposition are suffi ciently high, then both
bedding and burrows may be distinct (Barwis
1978
) .
Variation in seasonal bioturbation may be refl ected in
deposits as intercalated, laminated and burrowed beds.
The laminated beds characterize early winter when
bioturbation is low, whereas the bioturbated beds are
formed during summer when faunal activity is high
(in response to temperature and salinity variations,
which are commonly a response to fl uvial inputs).
A general model for tidal pointbar facies is illus-
trated in Fig.
11.16
(Barwis and Hayes
1979
). The