Geology Reference
In-Depth Information
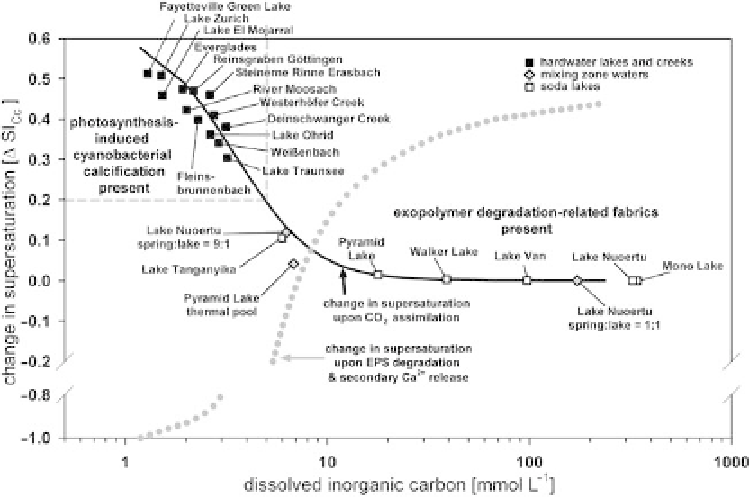
Fig. 16. Numerical simulations demonstrating the effect of photosynthetic CO
2
assimilation (black solid line) and
heterotrophic exopolymer degradation associated with secondary Ca
2þ
release (grey dotted line) on calcite saturation, as
dependent on the bulk phase dissolve inorganic carbon concentration. For details of calculation and assumptions see Arp
et al. (2001a, 2003).
environmental conditions (Riding 2000). Following
investigations of biofilm calcification in present-
day saline and freshwater settings and model cal-
culations using a given CO
2
removal and an
initial 10-fold calcite supersaturation (equivalent
to a 7-fold aragonite supersaturation), it has been
demonstrated that cyanobacterial photosynthesis
causes calcareous tubular microfossils to form
only in Ca
2þ
-rich and comparatively poorly pH-
buffered settings, (Fig. 16; Arp et al. 2001a). This
relation reflects the solubility product and pH
buffering by the dissolved inorganic carbon pool.
Consequently, calcareous cyanobacterial microfos-
sils can be used to trace secular changes in seawater
Ca
2þ
throughout the Phanerozoic when taking into
account palaeo-partial pressure curves for carbon
dioxide (Arp et al. 2001a). In turn, the enigmatic
lack of calcified cyanobacteria in stromatolite-
bearing Precambrian sequences can be explained
as a result of high dissolved inorganic carbon
concentrations (Arp et al. 2001a), alternatively to
other hypothesis (e.g. Riding 1982, 2006; Knoll
et al. 1993).
The results from studying tufa-forming biofilms
in karstwater streams support this view. Although it
was initially assumed that cyanobacterial calcite
tubes in tufa stromatolites reflect passive encrusta-
tion (inorganic mineralization) (e.g. Riding 1991:
32; Arp et al. 2001a: p. 1702), the present microsen-
sor data indicate that these tubes are directly formed
as a result of cyanobacterial photosynthetic activity
at comparatively low dissolved inorganic carbon
concentrations and low pH buffering.
On the other hand, the enormously diverse and
abundant non-phototrophic microbial community
apparently does not promote calcium carbonate pre-
cipitation in tufa-forming biofilms of karstwater
streams. Although detailed pathways of exopolymer
as well as low-molecular-weight substrate consump-
tion remain to be investigated, heterotrophic activity
of prokaryotes rather promotes the maintenance of
porosity in tufa biofilms, and possibly break-down
of inhibiting exopolymers at tufa stromatolites dis-
continuities. Here, overgrowth of existing calcite
crystals (neomorphism) in observed. Strikingly, the
non-phototrophic prokaryotes detected in the inves-
tigated tufa biofilms are almost exclusively aerobic.
Indeed, model calculations suggest that the pH
decreasing effect of CO
2
generation by far surpasses
the effect of ion activity product increase by second-
ary Ca
2þ
release from degraded exopolymers
(Fig. 16). This is due to the relatively low pH buffer-
ing in tufa stream waters when compared to soda lake
waters. In such lakes, exopolymer degradation and
secondary Ca
2þ
release effectively causes CaCO
3
precipitation because the simultaneously released
CO
2
is buffered by the high dissolved inorganic
carbon concentrations. The corresponding microbia-
lite fabrics, especially sickle-cell like shrinkage
voids or net-like calcified EPS structures, therefore
Search WWH ::

Custom Search