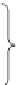Environmental Engineering Reference
In-Depth Information
rate of invasive populations and these stochastic factors can be accounted for in
integro-differential models (Lewis and Pacala 2000).
Boulant et al. (2009) developed a cellular automaton model (see Chap. 8) for the
spread of two pine species (
Pinus sylvestris
L. and
Pinus nigra
Arn. ssp.
nigra
)in
Mediterranean grasslands. This stochastic stage-structured integro-differential
model allows simulation of pine expansion within a 500m-wide rectangular gridded
landscape divided into 25
25 m cells. The demographic component of the model
accounts for the whole complexity of the pine life cycle: trees begin to produce
cones at the age of 10 years and cone production is age-(height-)dependent and
density-dependent. Therefore the life-cycle was divided into 15 stages (Fig.
16.4
):
nine seedlings age-classes (from 1-year old (S
1
) to 9-year old (S
9
)) and six adult
classes. Adults were divided into four 2-m height classes (A
2-4m
to A
>
8m
)of
increasing fecundity. When the total density of adults in a cell reaches the maximal
density of isolated trees (
D
is
), adults are considered as dominant in woodland
(A
dom
). Individuals reaching maturity when the total density of adults in a cell is
larger than the maximal density of dominant trees (
D
dom
) are considered as sup-
pressed (A
sup
), i.e. with a reduced fecundity. When the maximum tree density
(
D
max
) is reached, no more seedlings can establish. Seedling recruitment was
modelled explicitly as a function of tree fecundity and of the interactions between
shrub cover, post-dispersal seed predation, grazing pressure, grass competition, and
drought (see Boulant et al. 2009 for more details on demographic parameters).
The dispersal kernel of the pine model is a mixture of two exponential functions
that combines a small proportion (
p
) of long-distance dispersal and a larger
proportion (1 -
p
) of short-distance dispersal, which was estimated from field
data. Since dispersal was supposed to be isotropic, a one-dimensional form of the
kernel was used although the landscape was two-dimensional. The probability that
a seed produced in a cell
i
reaches a cell
j
located at a distance
x
ij
thus writes:
kx
ij
¼
p
2
e
2x
ij
=
a
1
p
a
1
2
e
2x
ij
=
a
2
p
a
2
þ
ð
1
p
Þ
, where
a
1
and
a
2
are the mean distances of
S
Immature stages (Seedlings)
A
su
p
A
dom
A
Mature stages (Adults)
D
A
>
D
dom
D
A
<
D
dom
2m-height classes
for isolated trees
s
j
= 0 if
D
A
>
D
max
D
A
>
D
is
Dominant
S
S
A
2-4 m
A
>8 m
Woodland
9
Suppressed
Fig. 16.4 Life cycle graph for the pine model [simplified from Boulant et al. (2009)]. Adult
density thresholds (in a 25
25 m cell): maximum density of isolated trees (
D
is
), maximum
density of dominant trees in woodland (
D
dom
), maximum tree density (
D
max
).
D
A
: adult density in a
cell (including isolated, dominant and suppressed trees), s
j
: seedling establishment rate