Chemistry Reference
In-Depth Information
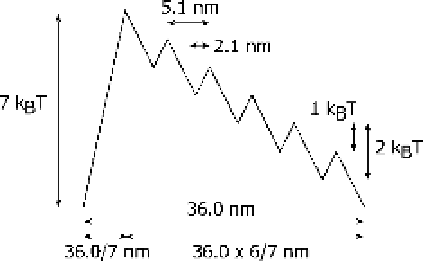
Figure 2.11 Computer simulation of multiple
head cooperative activity undergoing stochastic
steps [25]. We simulated the cooperative
behavior betweenmyosin heads under a periodic
and asymmetric potential as shown in
Figure 2.10b by numerically solving the Langevin
equation,
crossbridge; (2) the ratio of the potential rise to
decline was 1 to 6 and the depth of the potential
at the bottom was 2 k
B
T; (3) the pitch of the
potential and the average intervals of myosin
heads were 36 and 43 nm, respectively;
(4) the number of heads interactingwith the actin
filament was 11 (
20% overlap between actin
and myosin filaments); (5) the rotation angle of
the actin filament was 90
; and
(6) the rate constant (k
þ
) for the rebinding
of the heads to actin after the rewinding of the
actin filament was 100 s/head. The potential
slope was assumed to be smaller than that
estimated in the present experiment. The strain
exerted on the neck domain would be much
smaller during free shortening in muscle
because the head is tethered to the myosin
filament via a flexible
0
¼r
dx
i
/dt
dU(x
i
,t)/dx
þ
F(t)
A
i
,
where x
i
is the position of i-th myosin head;
r¼
10
3
pN/ns/nm is a drag coefficient;
F(t) is the random force obeying a Gaussian
white noise characterized by the ensemble
average,
8.8
<
F(t)
>¼
0 and
h
F(t)F(s)
i¼
2 k
B
T
r
d
s), where k
B
is the Boltzmann constant, T is
the absolute temperature, and k
B
T
(t
4.1 pN/nm;
A
i
is the interaction force between the
neighboring heads described as k(xi
i
¼
-helix (S2), while the head
is directly attached at its tail end to the probe in
the presentmeasurement system. Thus, because
the strain is smaller, so too is the potential slope.
(a) Schematic graph of the potential function,
U(x). (b) A typical time course of the movement
of an actin filament. (c) Histogram of the sliding
distance of actin filaments per ATP. The average
sliding distance of actin filaments was 58.4 nm
per ATP.
a
x
i1
)
k(x
i þ 1
x
i
), where k is the spring constant
connecting the heads. The potential slope along
the actin helical pitch was simplified to be a saw-
tooth shaped potential. The drag coefficient was
set to be larger than it is in solution so that the
velocity of the heads was equal to the maximum
velocity in Figure 2.6. Other parameters were
chosen such that (1) k was 0.1 pN/nm, which is
approximately one tenth as large as that of a rigor
man-made devices that operate at a much higher input energy level than k
B
T to avoid
the in
uence of thermal noise. For example, the input energy level of a transistor is
10
7
k
B
T in a
computer [77]. Szilard has shown by his model experiment of Brownian particles that
the minimum energy required for processing one bit data is k
B
Tln2
200 k
B
T and the energy used for processing just one bit data is
2
0.7 k
B
T [78].
Thus biological motors likely save energy by utilizing thermal
fluctuation. However,
the action of molecular motors driven by thermal
fluctuation is stochastic and
ambiguous. Although this stochastic and ambiguous nature is an impediment for
man-made machines, our computer simulations suggested that when stochastic
myosinmotors assemble to formsystems likemuscle, they show
exible and adaptive
¼