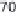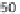Environmental Engineering Reference
In-Depth Information
and we have no idea what fraction of killed animals was
brought to camp sites or how many people it served.
Moreover, extreme spatial variability of the fragmentary
evidence permits no quantitative generalizations.
By the time anthropology was ready to study directly
the energetics of foraging societies (only well after WW
II), most such societies were either extinct or affected by
contact with neighboring pastoralists or farmers. Some
of the best records of relatively unchanged foraging sub-
sistence are available for the !Kung, 0Kade, and G/wi
groups of the Basarwa (Bushmen, the San), gatherers-
hunters of the late 1950s and early 1960s, just before
the rapid disappearance of this traditional way of life
(Lee 1979; Tanaka 1980; Silberbauer 1981), but they
pertain to a marginal environment and tell nothing about
the situation in more equable climates and fertile areas
where gathering and hunting were abandoned millennia
ago in favor of settled cultivation.
Systematic appraisals make clear that forager energetics
is a matter of peculiarities and exceptions rather than
of close similarities and general rules (Lee and De Vore
1968; Service 1979; Winterhalder and Smith l981; Kelly
1983; Price and Brown 1985; Kelly 1995; Gowdy 1998;
Lee and Daly 1999; Stanford and Bunn 2001; Panter-
Brick, Layton, and Rowley-Conwy 2001; Barnard 2004;
Frison 2004). Large differences in habitats and diets
translated into population densities differing by up to 2
OM. Minimum population densities of groups that
depended on mixtures of gathering and hunting activities
were on the order of 1/100 km
2
(tropical Aeta of the
Philippines, the Semang of Malaya, the boreal Micmac
of eastern Canada). The rates were 1 OM higher in sea-
sonally dry tropical environments (Kalahari Basarwa at
7-10/100 km
2
). Groups heavily dependent on fishing
had densities up to 1 OM higher (Pacific Northwest's
5.9 Approximate contributions of gathering, hunting, and
fishing to the diets of some foraging societies that survived into
the twentieth century. Plotted from data in Murdock (1967).
Nootka and Kwakiutl at about 60/100 km
2
, Makah at
nearly 90/100 km
2
).
Regardless of the prevailing source of food energy (fig.
5.9), large seasonal variations of staple food abundance,
as well as its annual fluctuations, resulted in highly irreg-
ular utilization patterns. Kelly (1983) suggested a cover-
age index (total exploited area/total residential mobility
distance) to indicate the intensity of land utilization. Pre-
dictably, it would be highest for gathering societies and
up to 1 OM lower for hunters. Primary and secondary
productivity and the shares of accessible edible biomass
are the key ecosystemic variables needed to evaluate for-
ager energetics. Forest phytomass is mostly in indigest-
ible lignin and cellulose of tree trunks; fruits and seeds
are a very small portion of the total, are commonly inac-
cessible in high canopies and are often well protected by
hard coats, requiring fairly energy-intensive processing
before consumption.