Agriculture Reference
In-Depth Information
a
b
Dy10 N Dx5
Dx5
Ax2*
Bx7
By9
Dy10
HMW-GS
c
Nontransformed
2.8× Dy10
2.6× D×5
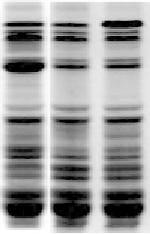
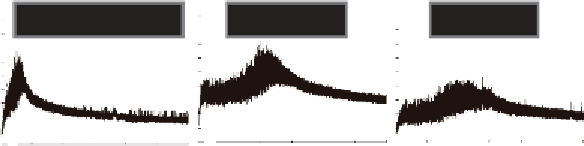
Fig. 18.3
Mixing characteristics of transgenic wheat fl ours with increases in either HMW-GS Dy10 or Dx5. (a) SDS-PAGE
of seed proteins of transgenic wheat lines containing increased levels of Dy10 (left lane) or Dx5 (right lane) compared with
their nontransformed parent (N). (b) Picture of 2-g mixograph instrument, showing the dough after development. The mixo-
graph is used to measure changes in resistance over time. (c) Comparison of mixograph traces for the three fl ours whose seed
proteins are shown in (a). Resistance in arbitrary units is plotted against time for 30 minutes. Increases in either Dy10 or Dx5
improve dough strength and mixing tolerance as shown by the thicker traces, longer times to peak resistance, and lower
slopes after peak resistance, compared with the nontransformed parent. However, the peak resistance achieved by mixing
dough with extra Dy10 is higher than the peak for the dough with extra Dx5, indicating that the effects of the two subunits
on dough development are different. (Photo courtesy of Frances Dupont.)
obtain transgenic plants with lower starch-
branching activities in their kernels. One trans-
genic plant had only 10% of the wild-type SBE
I activity; its seed starch had less crystalline
structure and gelatinized at a lower temperature
than starch from the nontransformed parental
seed. Regina et al. (2006) used RNAi constructs
to suppress expression of the genes encoding
SBEII a and/or b isoforms. Seeds from plants
with suppression of both isoforms contained
starch that was over 70% amylose, the linear
unbranched form that is more resistant to mam-
malian digestion. Rats fed wholemeal prepared
from transgenic seed exhibited improved indices
of large bowel function but the same growth rates
as rats fed meal from non-transgenic seed (Regina
et al., 2006).
Transformation has played a role in elucidat-
ing the genetic basis for grain hardness, a char-
acteristic that determines how much force is
needed to mill grain to fl our. The force require-
ment determines how much starch damage occurs
during milling, which is associated with water
absorption and bran contamination of white
fl our. Transformation experiments were used to
prove that the puroindoline genes
Pina-D1
and
Pinb-D1
together control grain hardness. Addi-
tion of
Pinb-D1a
(wild-type allele) under control
of a HMW-GS gene promoter to hard red spring
wheat 'HiLine', which contains a partially func-