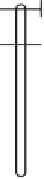Agriculture Reference
In-Depth Information
C/H S/T C/K
4B
4B
4B
0
barc193
Rht-B1
RGA13(2)
wPt-4699
12
0
P40/M60-8
0
16
wmc48c
4
P44/M54-7
gwm113
18
5
P46/M41-5
19
gwm495
11
wmc141
wg180a
gwm192B
20
wPt-1931
21
34
fba363
37
germin_B
bcd15
P41/M51
22
51
Rht-B1
56
P44/M54-6
23
gwm149
cdo669b
39
P32/M32-2
wmc48c
31
P36/M40
57
51
Rht-B1
34
wmc47
vutPDIb
wPt-1708
wmc48c
58
40
gwm6
56
62
gwm149
50
wmc349
ksuC2
61
s478tctg
63
gwm495
wPt-6209
P36/M40-9
P36/M36-6
P36/M40-2
P37/M35-5
63
68
ksuC2
66
77
gwm6
wmc47
70
gwm6
79
wmc47
83
wPt-0391
77
wPt-5996
92
101
P36/M37-1
Fig. 11.20
Collocation of leaf (TEL) and grain (TEG) transpiration effi ciency, and stomatal aperture trait QTL (SC
=
stomatal
conductance, CT
specifi c leaf area) and plant height (PH) QTL
for chromosome 4B measured in the Cranbrook/Halberd (C/H), Sunco/Tasman (S/T), and CD87/Katepwa (C/K) populations;
TEG was determined for the S/T population only.
=
canopy temperature, SPAD
=
chlorophyll content, SLA
=
contributed high-CID alleles for CID measured
on mature grain (Fig. 11.20). This inconsistency
may account for the low genetic correlation for
leaf and grain CID (
r
g
= 0.24,
n
= 159). This change
in allelic state highlights the need to consider vari-
ation in developmental traits and the potential for
confounding environmental factors with yield
when measuring CID on grain. Further, it high-
lights the importance of controlling variation in
development when indirectly selecting for yield
via leaf or grain CID (
cf
. Ferrio et al., 2007).
The expense of measuring CID with a mass
spectrometer (approximately US$40 per sample)
underscores the need to identify surrogate traits
for screening large populations. Colocation of
CID QTLs with QTLs for stomatal-related traits
gives some indication of the underlying physio-
logical basis for variation in TE in wheat. For
example, the 4BS low-CID QTL was associated
with increased chlorophyll content, greater sto-
matal conductance, and cooler canopy tempera-
tures (Fig. 11.20). This suggests the potential for
indirect selection for CID using an infrared ther-
mometer or SPAD chlorophyll meter, perhaps as
early-generation selection on head rows. Use of
NIR also offers the potential for inexpensive
assessment of CID in large segregating wheat
populations (Fig. 11.6).
biomass and yield. Maintenance of excessive leaf
area when water is limiting can contribute to
unnecessary water loss and sometimes lead to
haying-off and reduced grain yields (Fischer
1979; van Herwaarden et al., 1998). On the other
hand, reductions in leaf area, particularly around
anthesis and throughout grain fi lling, can reduce
assimilation and accumulation of carbon neces-
sary for grain set and grain fi lling. The impor-
tance of extending the duration of green leaf area
as a means of maintaining postfl owering photo-
synthesis and supply of assimilated carbon has
been widely recognized (Richards 2000; Foulkes
et al., 2004). Between 50% and 90% of fi nal grain
weight and yield is derived from postanthesis
assimilation (Bidinger et al., 1977; Schynder
1993). Loss of leaf area and subsequent reduc-
tions in grain yield have been reported in wheat
(Gelang et al., 2000), while daily delays in fl ag leaf
senescence were associated with an estimated
0.17 t ha
−1
increase in grain yield and 2.7 kg ha
−1
increase in nitrogen content (Gooding 2007).
A common response of water-stressed plants is
accelerated and premature leaf senescence, leading
to a reduction in the amount of photosynthetic
leaf tissue (Sirault et al., 2004). Genotypic
increases in leaf area duration can be achieved via
selection for genes that delay the onset and rate
of leaf senescence (termed stay-green) (Thomas
and Howarth 2000) or for genes that protect the
leaf from stresses affecting leaf senescence (e.g.,
disease, heat, and water stress). Genotypic varia-
tion in the ability to maintain leaf area under
Maintenance of leaf area
A growing wheat crop must accumulate and then
maintain leaf area toward development of fi nal