Agriculture Reference
In-Depth Information
Early leaf area development
1,200
-
tin
0 kg N
-
tin
160 kg N
+
tin
0 kg N
+
tin
160 kg N
1,000
In Mediterranean-type environments, crops are
typically sown on the fi rst rains, and water for
growth is supplied as current rainfall. Here, faster
leaf area development should reduce soil evapora-
tion to increase crop water-use effi ciency, yield
(López-Castañeda and Richards 1994), and com-
petitiveness with weeds (Coleman et al., 2001).
Greater early vigor may also benefi t crops through
increased root growth early in the season
(Fig. 11.9 and 11.10; Palta et al., 2007), while
improved light interception should increase crop
growth rate, biomass, and grain yield for late-
sown wheat crops or in environments where
crop duration is shorter (Regan et al., 1997).
Movement toward environmental sustainability
and concerns over increasing diesel prices have
increased interest in conservation farming and
especially reduced tillage. However, hard soils
and retained stubble common to conservation
farming slow early growth, reducing crop biomass.
Greater intrinsic early vigor has potential to
overcome constraints imposed by reduced tillage
(Watt et al., 2005).
Genotypic increases in early vigor have been
shown to be associated with greater biomass and
grain yield for wheat grown in Mediterranean
environments (Whan et al., 1991; Botwright et al.,
2002). Despite the potential benefi ts for greater
early vigor in cereals, there has been little evi-
dence of targeted breeding for this trait, outside
of efforts to improve stand establishment and
promote vegetative canopy closure in early-
planted, dual-purpose management systems
(Carver et al., 2001). This may partly refl ect low
heritability and reduced selection response for
seedling leaf area and biomass (Rebetzke and
Richards 1999). Numerous morphological factors
with moderate to high heritability contribute to
increased early vigor in wheat. Variation in early
vigor is associated with differences in rate of seed-
ling emergence, kernel and embryo size, coleop-
tile tiller size, and specifi c leaf area of seedling
leaves (López-Castañeda et al., 1996; Rebetzke
and Richards 1999; Rebetzke et al., 2004).
Development of wheat cultivars with the capac-
ity to emerge from deep sowing (110 mm) would
800
600
400
200
0
20
40
60
80
100
120 140
160 180 200
Days after sowing
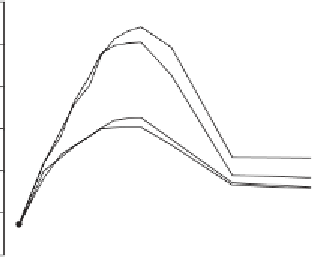
Fig. 11.16
Change in shoot number over time for sister lines
with (
) the tiller inhibition (
tin
) gene
when supplied with starting fertilizer and then either 0 or
160 kg ha
−1
additional fertilizer (Duggan et al., 2005).
+
) and without (
−
production of wheat pasture and grain in a single
cropping season.
De-tillering studies by Jones and Kirby (1977)
and Islam and Sedgley (1981) indicate the poten-
tial for increasing grain yield through reductions
in the number of nonsurviving shoots. The ability
to genetically control tiller number has major
potential in managing target tiller and spike
number for a given environment or management
regime. Genotypic variation exists to modify tiller
number. For example, simply reducing the period
from sowing to terminal spikelet to hasten fl ower-
ing can reduce tiller number (Rebetzke et al.,
2008b). A major tiller inhibition gene (
tin
) has
been located linked to glume pubescence (
Hg
)
and the microsatellite marker
Xgmw136
on chro-
mosome 1AS (Spielmeyer and Richards 2004).
This gene reduces potential tiller number from as
many as 15 to between 1 and 5 tillers per plant
depending on genetic background (Fig. 11.16)
(Hendriks 2004; Duggan et al., 2005). Under
water-limited conditions, near-isogenic lines con-
taining the
tin
gene produced greater WSC,
increasing kernel size and reducing the propor-
tion of shriveled kernels (Mitchell et al., 2006).
Further, lines containing the
tin
gene increased
partitioning of carbon to roots, increasing root
biomass (Hendriks 2004; Fig. 11.11).