Agriculture Reference
In-Depth Information
postanthesis assimilation (Bidinger et al., 1977).
However, conditions in rainfed environments are
rarely favorable as drought (and disease) can
inhibit photosynthesis and/or decrease leaf area
to reduce carbon supply to developing grain.
This reduced carbon supply commonly results
in lower grain yields and the production of
small and shrivelled kernels that devalue the crop
and reduce returns to growers (Ruuska et al.,
2006).
Assimilated carbon can accumulate temporarily
in the stem and leaf sheath during the later growth
stages of cereals. This nonstructural carbohydrate
pool accumulates prior to, during, and after
anthesis, and can be later remobilized and
transported to developing grain (Schnyder 1993).
In wheat, this stem reserve carbohydrate is stored
in the form of WSC, which principally consists of
fructosyl-oligosaccaharides (fructan) (Ruuska
et al., 2006). Total WSC may attain levels of more
than 40% of total stem dry weight in wheat,
and its remobilization can make a signifi cant
contribution to fi nal grain yield and kernel size
(Schnyder 1993 and references therein). This
contribution varies from season to season and can
be as high as 30% to 50% of total grain weight
when conditions are unfavorable for photosynthesis
and between 10% and 20% when conditions are
favorable (Bidinger et al., 1977; Schnyder 1993;
van Herwaarden et al., 1998).
The importance of stem WSC reserves under
drought-stress conditions has been demonstrated
in the fi eld (van Herwaarden et al., 1998; Foulkes
et al., 2002; Yang et al., 2007; Rebetzke et al.,
2008b) and predicted in crop modeling studies
(Asseng and van Herwaarden 2003). Genotypic
variation for WSC is large and repeatable across
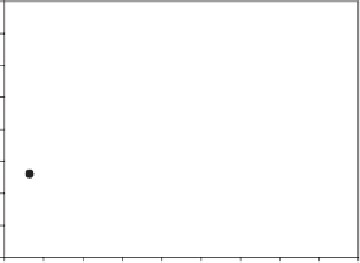
diverse environments in wheat (Fig. 11.12)
(Ruuska et al., 2006). Breeding-era studies have
demonstrated increases in WSC concentration
with release of new wheat cultivars for some
Australian wheat breeding programs (e.g.,
Western Australia in Fig. 11.13) and at CIMMYT
in Mexico (Fig. 11.14) (van Herwaarden and
Richards 2002). However, no relationship was
observed for release of wheat cultivars in eastern
Australian breeding programs (Fig. 11.13).
Release of higher-yielding, UK wheat cultivars
320
300
H45
Westonia
280
QG225
260
Cranbrook
Tasman
Quarrion
CD87
240
Janz
Halberd
Egret
220
Silverstar
Chara
Sunco
Sunstate
200
Sunstar
180
160
140 160
180
200
220
240 260
280 300
320
WSC concentration - droughted (mg/g DW)
Fig. 11.12
Relationship of mean water-soluble carbohy-
drate (WSC) concentration at early grain-fi ll for different
Australian wheat genotypes sampled at water-limited and
irrigated sites (Y
=
86
+
0.73X,
r
2
=
0.74) (Rebetzke et al.,
2008b).
from 1972 to 1995 was associated with increasing
WSC concentration (Shearman et al., 2005). The
environment in Western Australia is typically
Mediterranean, so that the crop grows on current
rainfall. It would be advantageous in this
environment to develop large anthesis biomass
with the potential to remobilize a large portion of
this dry matter to grain (Fischer 1979).
In studies undertaken across multiple
environments, Rebetzke et al. (2008b) observed
positive associations (
r
g
= 0.27-0.43) between
WSC concentration and grain yield, and between
WSC concentration and kernel size (
r
g
= 0.60-
0.62), in three unrelated wheat populations. Lines
high for WSC concentration (milligrams WSC
per gram dry weight) commonly were earlier for
fl owering, were reduced in plant stature, and
produced fewer tillers per plant, leading to
reduced anthesis and fi nal plant biomass (Rebetzke
et al., 2008b). Nitrogen content was also lower for
high-WSC lines, consistent with high WSC
measured in wheat grown with low fertilizer
nitrogen (van Herwaarden et al., 1998). Lines
actually selected for high WSC content (i.e.,
WSC per ground area, or g m
−2
) produced larger
kernel size and higher grain yields but main-
tained higher tiller numbers, leading to increased