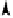Biology Reference
In-Depth Information
Exon 4 Exon 5
Exon 6
cDNA
STOP
4
5
6
Alu
HLA-DR-
1 gene
Extended
exon 5
Exon 4
Exon 6
STOP
Alu
-containing cDNAs
Exon 4
Extended exon 5
STOP
Figure. 8.9.
A resident intronic
Alu
sequence is incorporated into exon 4 of the HLA-DR-
β 1 mRNA by intron sliding (from Labuda
et al
., 1995). Among three variants of the
HLA-DR-β 1 cDNA, detected by library screening, the upper one is considered to be the
usual form, whilst the two others are alternatively spliced, apparently due to a lack of
splicing at the intron 5 donor site. As a result, exon 5 is extended into a nearby
downstream
Alu
sequence in intron 5 either to include a stop codon within the
Alu
sequence or to be spliced with exon 6 (the open reading frame in the extended exon 5
matches that of exon 6). These three cDNA clones may illustrate two phases of 'intron
sliding': inactivation of an existing splice site followed by activation of a cryptic one.
8.5 Gross gene duplications in evolution
Gene duplication (or partial duplication) events are a fairly uncommon cause of
human genetic disease (reviewed by Hu and Worton, 1992; Mazzarella and
Schlessinger, 1998). Two distinct mechanisms are currently envisaged: (i)
homologous
unequal recombination
either between homologous chromosomes or sister chro-
matids and (ii)
nonhomologous recombination
at sites with minimal homology.
Topoisomerase cleavage sites have been reported to be associated with pathologi-
cal gene duplications (Kornreich
et al
., 1990; Hu
et al
., 1991) and potential sites
for topoisomerases I and/or II have been found to coincide with the breakpoints
of duplications in the human factor VIII (
F8C
; Xq28; Casula
et al
., 1990) and dys-
trophin (
DMD
; Xp21.2-p21.3; Hu
et al
., 1991) genes. These observations are
potentially interesting since topoisomerase activity has been implicated in several
cases of nonhomologous recombination (Bullock
et al
., 1985).
One of the best studied gross duplications in human genome pathology is the
1.5 Mb duplication of the short arm of chromosome 17 associated with Charcot-
Marie-Tooth disease type 1A. This recurring duplication is thought to be medi-
ated by homologous unequal recombination between two misaligned ~30 kb
CMT1A-REP repeat sequences flanking the CMT1A region in direct tandem ori-
entation (Reiter
et al
., 1996). In humans, these repeats are 98% identical.
Chimpanzees have two copies of a CMT1A-REP-like sequence, whereas gorilla,