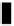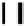Biology Reference
In-Depth Information
0
2
4
6 kb
p
p
p
1
(L1)
23
(M)
4
(L2)
56
78
9101112
Figure 5.3.
Exon/intron distribution in the human aldolase A (
ALDOA
) gene. Exons 1,
3, and 4 correspond to leader exons L1, M2, and L2. The positions of the three
alternative promoters are denoted by p (redrawn from Mukai
et al
., 1991).
identified by
in vitro
studies are actually always utilized
in vivo
. White
et al
. (1998)
identified the
in vivo
transcriptional initiation sites of the cystic fibrosis trans-
membrane conductance regulator (
CFTR
; 7q31.3) gene in both human and
mouse. Tissue-specific variation in the position of the transcriptional initiation
sites was noted in both species but the sites were not conserved between equiva-
lent tissues. This finding suggests that the precise mechanism of transcriptional
initiation for a given gene may not be absolutely conserved between species.
5.1.8 Developmental changes in gene expression
In higher primates (both platyrrhine and catarrhine), the
-globin (
HBG1
,
HBG2
; 11p15.5) genes are expressed during fetal life whereas in nonprimates and
prosimians, the genes are expressed in the embryo. The conversion of the
HBG1
and
HBG2
genes to a fetal pattern of expression must therefore have occurred
after the divergence of simians from prosimians some 55 Myrs ago. Implicit in
this conversion is the activation of the
-globin gene in fetal life (a stage at which
it was previously repressed) and repression of the
-globin gene in embryonic life
at which stage it was previously active. Prosimians are characterized by the pos-
session of only one
-globin gene whereas higher primates possess two copies. The
promoter of one of the duplicated
-globin genes may thus have been able to
escape the influence of natural selection and in so doing accumulate mutations
that served to alter the timing of its developmental expression. Any beneficial
changes thus acquired could then have been readily transferred to the other
-glo-
bin gene by gene conversion.
A burst of sequence change occurred after the divergence of simians from
prosimians but before the divergence of Old World monkeys from New World
monkeys (Fitch
et al
., 1990, 1991). Most of these changes were then conserved
during the subsequent evolution of the simian
-
globin gene promoter in transgenic mice have shown that all the sequence
changes necessary for changing to the fetal pattern of gene expression are located
within a 4 kb fragment containing the
-globin genes. Studies of the
-globin gene (TomHon
et al
., 1997).
However, within 260 bp of the transcriptional initiation site, there are 19
nucleotide changes specific to simians, 16 of which are located in or near highly
conserved sequence motifs (Chiu
et al
., 1997; Fitch
et al
., 1990; 1991). Further,
comparison of the human and galago (a prosimian) sequences reveals 57
nucleotide differences over the same region with the majority again being located