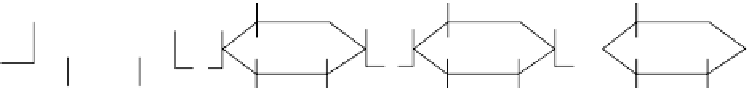Environmental Engineering Reference
In-Depth Information
(a)
α
-(1
→
4) linkage
CH
2
OH
CH
2
OH
CH
2
OH
CH
2
OH
O
O
O
O
H
2
OH
OH
OH
OH
OH
O
O
HO
OH
OH
OH
OH
(b)
α
-(1
→
6) linkage
CH
2
OH
CH
2
OH
CH
2
OH
CH
2
O
O
O
O
OH
OH
OH
OH
O
O
O
O
OH
OH
OH
OH
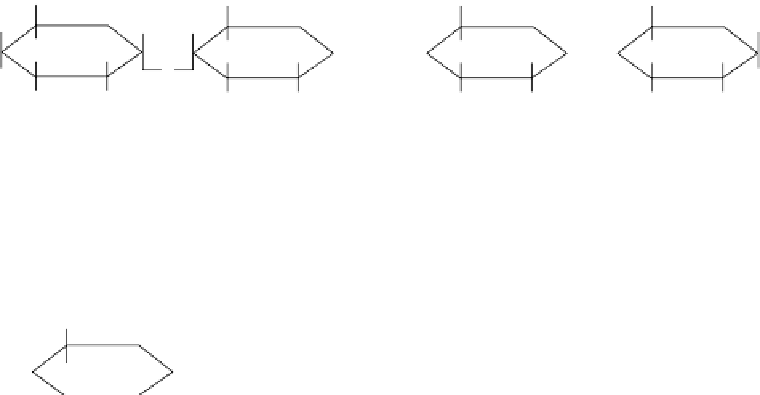
FIGure 31.1
Structure of amylose (a) and amylopectin (b) showing α-(1,4) and β-(1,6) linkages. (Adapted
from Nowjee N.C.,
Melt Processing and Foaming of Starch
, University of Cambridge, 2004. available at
http://www.cheng.cam.ac.uk/research/ groups/polymer/RMP/nitin/Foaming.html (accessed June 2010).)
The extent of crystallinity of native starch granules ranges from approximately 15% for high
amylose starches to approximately 45-50% for waxy starches (Copeland et al. 2009). Amylopectin
chains with more than ten glucose units are organized into double helices, which are arranged into
either A- or B- crystalline forms that may be identified by characteristic x-ray diffraction spectral
patterns. The double helical structures within the A- and B-type crystalline forms are essentially
the same (Gidley 1987; Imberty et al. 1991). However, the packing of the helices in the A-type
crystalline structure is more compact than in the B-type structure, which has a more compact
structure with a hydrated core. The A-type crystal pattern has amylopectin molecules with shorter
chains (Jane 2006). Tuber starches and amylose-rich starches yield the B-type pattern, although
both types could occur (Buléon et al. 1998). Starch varies greatly in form and functionality between
and within botanical species and even from the same cultivar grown under different conditions,
hence providing starches of diverse properties (Copeland et al. 2009).
The susceptibility of starch to hydrolysis by α-amylase has been shown to vary with botanical
origin (Srichuwong et al. 2005). Starch digestibility is not necessarily related to total starch
content because the amylose/amylopectin ratio and processing affect the extent of starch digestion
(Reynolds et al. 1997). Although a minor component by weight, lipids can have a significant role in
determining the properties of starch. The lipid of native starches is highly correlated with amylose
content: the higher the amylose content, the higher the lipid present. Inclusion complexes with
lipids form mainly with the amylose component of the starch; hence, the amylose/amylopectin
ratio is an important factor that produces variability in the ability of natural starches to bind
lipids (Copeland et al. 2009). Because of its high degree of branching, the lipid binding capability
of amylopectin is considered to be much weaker than amylose. There is little direct evidence to
suggest that amylopectin forms true inclusion complexes with lipids, although some studies have
led to proposals that favor the interaction of some lipids and surfactants with outer amylopectin
branches (Hahn and Hood 1987; Eliasson 1994; Villwock et al. 1999). Complexes between amylose
and lipids can significantly modify the properties and functionality of starch. For example, these