Information Technology Reference
In-Depth Information
B
Extended-VITE
T
1
GO
T
2
V
1
V
2
GV
1
GV
2
P
A
1
A
2
Extended-FLETE
I
1
I
2
M
2
S
2
D
2
D
1
S
1
M
1
R
1
R
2
X
1
X
2
spindle response
spindle response
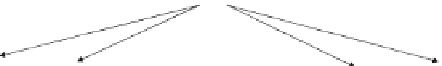
Fig. 10.1: (continued)
cells, the tonic cells, the reciprocal cells, and the bidirectional cells known to play a
role in voluntary movement initiation and control [19, 21].
Third, they failed to identify the site of origin of the triphasic pattern of muscle
activation [2]. Is the triphasic pattern cortically or subcortically originated [2]? If
cortically originated, are the agonist and antagonist bursts generated from experi-
mentally identified cortical cell types? Does the afferent feedback from the muscle
spindles to the spinal cord play any role in maintenance of this pattern? Does the
feedback from the muscle spindles to the cortex play a role in the generation of the
second agonist burst?
These limitations were addressed successfully by the extended VITE-FLETE
with dopamine models of Cutsuridis and Perantonis [18] and Cutsuridis [15, 16, 17].
These models have answered issues concerning voluntary movement and proprio-
ception in normal and Parkinsonian conditions. The temporal development of these
models in normal conditions (i.e., without dopamine) is reviewed in detail in the
next section.
10.3 The Extended VITE-FLETE Models Without Dopamine
Figures 10.1a, b depict the extended VITE-FLETE models without dopamine of
voluntary movement and proprioception [15, 16, 17, 18]. Both models were based