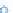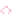Geology Reference
In-Depth Information
that using the same method and the uppermost
Belize curve as a datum, Gischler found that reef
accretion increased signifi cantly with depth, and
intervals dominated by massive corals accreted
faster than those dominated by branching acropo-
rids (E. Gischler, personal communication, 2006).
A fi nal possibility relates to the way in which
the data were collected. Existing models are
more often based on generalized patterns seen
throughout a core or along an entire reef over its
lifetime. In contrast, the measurements reported
here are from discrete intervals within single
cores. Furthermore, the data do not include
intervals with a mixture of branching and massive
corals or a predominance of detritus. This approach
was chosen to (1) allow for a more accurate char-
acterization of average palaeowater depth across
a single accretionary interval, and (2) address the
presumption that branching-coral reefs in shallow
water will accrete much faster than their massive
counterparts at depth. What effect might this have
on the data, and would it be suffi cient to explain
the difference between the expected depth- and
species-related patterns and those seen here?
Reefs do not accrete evenly, either spatially
or temporally. Using each interval between
dated samples adds variability and undoubt-
edly contributed to the high variance seen in
Fig. 8. However, when the patterns from a par-
ticular reef system were examined (Fig. 10a),
wholesale accretion rates between coral types
and with increasing depth remained similar, both
to one another and to the average rates for smaller
intervals reported above. For example,
A. pal-
mata
reefs from the shelf edge off both St. Croix
and Puerto Rico accreted at rates similar to those
from massive-coral reefs in deeper water, at the
same place and time. Similarly, branching- and
massive-coral reefs near Buck Island (St. Croix)
accreted at similar rates, regardless of the dom-
inant coral type. Whole cores showed the same
pattern; cores from the same time frame within
a single reef yielded accretion rates similar to
each other and to the averages reported above
for smaller intervals regardless of depth or facies
(Fig. 10b).
(a)
0
Coral samples
5
10
15
20
25
30
35
(b)
0
Reef accretion
BI-02
5
BI-04
Ap
10
PAR-02/03
LB-02
LB-03
15
PAR-19
LB-04
20
25
30
35
11
10
9
8
7
6
5
4
3
2
1
0
Age (Cal
14
C kyr BP)
Fig. 10.
(a) Ages versus water depths of all samples
examined for this synopsis. The coloured envelopes
delineate samples grouped by species for six reefs on
St. Croix and Puerto Rico. The general trend of each enve-
lope is interpreted as the overall rate of reef building by
that species in a single reef. Earlier reefs accreted at the
same rate regardless of species (fi lled =
Acropora palmata
;
open = massive) or water depth. Similar parity existed
in later reefs, except they built at slower rates. (b) Reef-
accretion pathways for selected cores examined for this
study. Note that, with the exception of the massive-coral
interval constrained by accommodation space (BI-04)
and one deep reef (PAR-19), accretion rates were similar.
Acropora palmata
intervals are shown by solid lines;
massive intervals are dashed. LB = Lang Bank (E St. Croix,
US Virgin Islands); BI = Buck Island (N St. Croix),
PAR = La Parguera (SW Puerto Rico). The shaded blue
band corresponds to the mean accretion rate for this
study (3.45 m kyr
1
).
(1980) concluded that, 'the internal structure of
the reef is controlled not only by the nature of the
dominant builders but also by the type and rates
of bioerosion'. Chazottes
et al
. (1995) proposed
that the greatest substrate loss off Morea was
due to grazing fi sh and urchins (2.33 kg m
2
yr
1
),
followed by macroborers (e.g. molluscs, worms:
0.20 kg m
2
yr
1
) and microborers (algae, fungi:
0.009: 2.33 kg m
2
y
1
). A similar hierarchy has
been proposed for the Great Barrier Reef (Kiene &
The possible role of bioerosion
References to biological destruction of substrate
date at least to Grant's 1825 description of tem-
perate species of
Cliona
, and were more formally
recognized when Neumann (1966) introduced the
term 'bioerosion'. More recently, Scoffi n
et al
.