Geology Reference
In-Depth Information
Accretion rate
Reef-accretion rates have typically been reported
for whole cores or reefs (for an excellent sum-
mary, see Dullo, 2005). In this paper, accretion
rates from individual intervals (i.e. between two
adjacent samples) within cores were chosen
over those for whole cores or facies, because palaeo-
water depth could be more accurately assigned
to a shorter time step. Intervals that contained
largely mixed coral types or were dominated by
detrital deposition were not included. Samples
that plotted above sea level or resulted in negative
accretion rates (i.e. the lower sample was younger
than the upper one) were not used. Finally, rates
calculated from closely spaced samples with age
differences less than the reported analytical errors
were also discarded. Of the 11 discarded accretion
calculations, all were faster than 10 m kyr
1
. All but
three were from palaeowater depths greater than
5 m. Futhermore, these shorter-term rapid rates
were invariably contained within longer intervals
that refl ected much lower net accretion rates.
Separate analyses of reef accretion were run
for (A) all corals regardless of species, and (B) a
comparison of
A. palmata
versus massive-coral
facies. The dominant corals within an interval
were determined by examining core logs. The
11 data points from abandoned
A. palmata
reefs
off Barbados (Fairbanks, 1989; Bard
et al
., 1990)
were used for the comparison between accretion
rates in massive and branching facies only. They
were eliminated from the depth-related compar-
ison because no independent verifi cation exists
for sea level at the time these deeper reefs were
active. All the Belize corals above the curve used
in this paper (Fig. 5) occurred within mixed-coral
intervals that violated the acceptance criteria.
Thus, all the data used from the cores of Gischler
& Hudson (2004) fell below the curve.
Atlantic Ocean
N
Caribbean Sea
1000 km
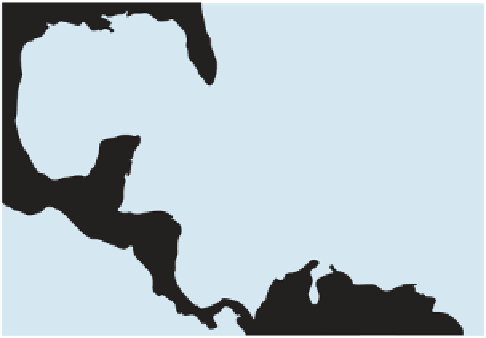
Fig. 6.
Map showing the locations of the sites from which
core data were derived (open circles).
Table 1.
Sources of core data used in this study.
Florida Lighty
et al
. (1982), Toscano & Lundberg
(1998). Toscano & Macintyre (2003)
Bahamas Lighty
et al
. (1982)
Martinique Lighty
et al
. (1982)
Puerto Rico Hubbard
et al
. (1997, unpublished data),
Lighty
et al
. (1982)
Antigua
Macintyre
et al
. (1985)
St. Croix
Adey
et al
. (1977), Burke
et al
. (1989),
Hubbard
et al
. (1990), Hubbard
et al
. (2005),
Hubbard (unpublished data), Lighty
et al
.
(1982), Macintyre & Adey (1990)
Barbados
Fairbanks (1989)
Panama
Macintyre & Glynn (1976)
Belize
Gischler & Hudson (2004)
fringing reefs to submerged but still active
shelf-edge reefs (e.g. Lang Bank and SW Puerto
Rico) and abandoned reefs that are now below the
depth where accretion can occur (e.g. Barbados).
The data are tabulated in Appendix 1.
The majority of the data points fell within
the upper 15 m of the water column (Fig. 7). As
expected, the fastest accretion rates were derived
from core intervals in water <10 m deep at the
time of deposition. However, accelerated accre-
tion was evenly split between branching- and
massive-coral intervals, and any depth-related
coral affi nities were weak above 15 m.
A. palmata
clearly thrived at depths much greater than is
generally accepted as its maximum, commonly
deeper than 10 m and occasionally down to 20 m.
More important, they were capable of signifi cant
accretion well below the 5-m depth limit pro-
posed by Lighty
et al.
(1982) and Macintyre (1988),
among others.
RESULTS
A total of 151 core intervals met the criteria
described above. Of these, 79 were from branch-
ing-coral facies, compared with 72 from massive-
coral intervals. Samples were recovered from
11 Caribbean and Atlantic locations (and from
multiple reefs at many of these), ranging from cen-
tral Florida in the north to Panama and Belize in
the southwest and Antigua/Barbados in the east-
ernmost Caribbean (Fig. 6; Table 1). Present-day
environments ranged from emergent barrier and