Agriculture Reference
In-Depth Information
PIN 1
GN
MP
BDL
PIN 4
GN
MP
BDL
PIN 7
B
A
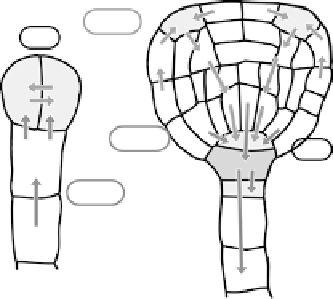
Figure 1.5
Auxin transport and distribution during embryogenesis. Sites of auxin accumulation are
shadowed. Arrows indicate routes of auxin efflux mediated by PIN1, PIN4 and PIN7. Also depicted are
proteins involved in embryo patterning and related to auxin transport (encircled) or auxin response. (A)
Two cell stage embryo - apical cell specification. Auxin accumulates in the proembryo through
PIN7-dependent transport via the suspensor. Auxin response (
mp
,
bdl
) and transport (
gn
) mutants show
defects in the establishment of the apical cell. (B) Triangular stage - from early globular stage on auxin
accumulates, in a PIN1- and PIN4-dependent manner, in the hypophysis, which is specified and is
further transported through the suspensor via a PIN7-dependent route. New sites of auxin accumulation
emerge at the tips of forming cotyledons.
mp
,
bdl
and
gn
show defects in root pole as well as cotyledon
establishment. Adapted from Friml
et al
. (2003), with permission.
1.5.3 Postembryonic organ formation
Embryo development establishes the basic body plan of both animals and plants.
However, the adult form of a plant also depends largely on postembryonic
development. Plants, unlike animals, can postembryonically initiate new organs
such as leaves, flowers, flower organs, ovules and lateral roots. The regular initiation
pattern of leaves and flowers (called
phyllotaxis
)isthe major determinant of adult
plant architecture. During organ formation, first a site of primordium initiation is
selected and then a new growth axis of the organ primordium is established. It seems
that PIN-dependent redirection of auxin flow and local accumulation of auxin play
a fundamental role in both of these processes. Exogenous auxin application is suf-
ficient to trigger leaf or flower formation in the shoot apex (Reinhardt
et al
., 2000)
or lateral root initiation (Laskowski
et al
., 1995), and endogenous accumulation of
auxin and its response was detected at the initiation site of incipient organs in shoots
and roots (Benkova
et al
., 2003). On the other hand, interference with PAT (AEIs,
pin
mutants) or auxin response (mp,
solitary
root) blocks organ formation (Okada
et al
., 1991; Przemeck
et al
., 1996; Fukaki
et al.
, 2002). In the shoot, PIN1 local-
ization in the outermost layer (L1) of the meristem undergoes dynamic rearrange-
ment toward these loci of auxin accumulation (Reinhardt
et al
., 2003). The pattern
of auxin accumulation and PIN localization suggests that auxin is transported to-
ward the meristem through the L1 cell layer. There, auxin becomes absorbed by