Agriculture Reference
In-Depth Information
A: SLA
B: HSP
C: PLC50
C.gloriosus
C.crassifolius
subgenus
Cerastes
C.cuneatus
C.maritimus
C.prostratus
C.megacarpus
C.leucodermis
C.oliganthus
C.hearstiorum
subgenus
C.spinosus
Ceanothus
C.cordulatus
C.griseus
C.integerrimus
0.75
1
1.25
0.5
1
6
8
10
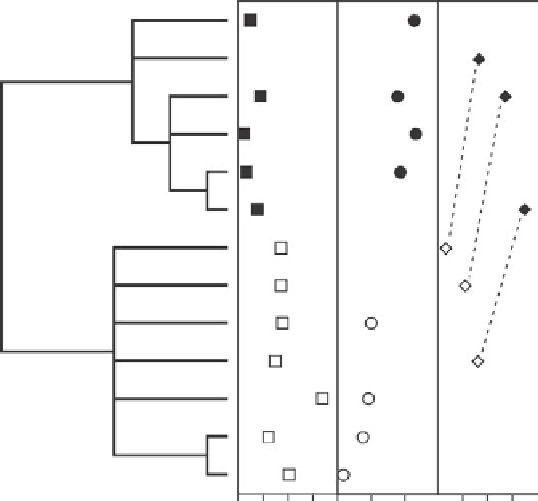
Fig. 9.12
Partial phylogeny of
Ceanothus
species illustrating divergence in regeneration strategy
and other functional traits. A: specific leaf area (log (mm
2
mg
1
)), B: heat shock proteins, C:
drought tolerance (
C
50% survival
MPa) and dashed lines connecting sympatric species pairs along
an altitudinal gradient. (From Ackerly 2003.)
California where different recruitment patterns combine within communities in
non-random associations (Keeley & van Mantgem
2008
).
Many of the postfire seeder genera are quite diverse (see
Table 3.4
) and this
phenotypic clustering of seeders is considered an example of evolutionary
conservatism when taxa have radiated into environments with very similar fire
regimes. This of course is a function of scale, as at the biome level conservatism
appears to be the norm in plant evolution (Crisp
et al.
2009
). This can be detected
by a higher mean relatedness (lower mean phylogenetic distance) between all
pairwise species of a fire-prone community than the one expected under a null
model (
Fig. 9.13
), and also higher than in a community where such a strong
filtering process as fire is not present (Verdu´ & Pausas
2007
).
Niche conservatism is evident in the North American
Ceanothus
where the very
diverse
Cerastes
lineage comprises only obligate seeding species that have radiated
into different crown fire shrubland communities in California, Arizona and east-
ern Mexico (Keeley
2000
). However, another chaparral genus,
Arctostaphylos
,
illustrates a pattern more consistent with an adaptive evolution model in that
10 of the 16 resprouting species have subspecies that are obligate seeders, reflect-
ing a close tracking of changing fire regimes: the resprouting populations are