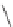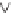Agriculture Reference
In-Depth Information
Infertile soils
Large and
high-nutrient
seeds
Intense seed predation
Canopy seed
storage
Myrmecochory
(-) Limit seedbank size
(-) Increase carbon cost for protection
(-) Less resilient to long fire cycles
(+) Reduce predation
(-) High carbon cost for “bribe”
(+) Reduce predation
(+) Disperse to open sites
Fig. 9.7
Hypothesized effects of nutrient-poor soils in fire-prone ecosystems on serotiny and
myrmecochory in Australian and South African MTC ecosystems.
Perhaps the greatest weakness of the rainfall reliability theory is that serotiny in
some lineages appears to be Tertiary in origin with important speciation events
occurring over the last 10 Ma (see
Chapter 10
). However, the subtle differences in
rainfall predictability between the northern and southern hemisphere MTCs have
likely fluctuated greatly over that period of time, in concert with Milankovitch
cycles and other longer-term changes (Bennett
1990
,
2004
).
One factor that has persisted through much of the Tertiary and ties together
centers of serotiny in Australian heathlands and South African fynbos is the
widespread occurrence of highly weathered nutrient-poor soils (see
Fig. 1.5
), and
thus nutrient-poor fire-prone vegetation (
Fig. 9.7
). There may be a link between
soils and serotiny since the oligotrophic soils in heath and fynbos put a premium
on nutrients for seeds (Stock
et al.
1990
; Vaughton & Ramsey
2001
; Groom &
Lamont
2010
). Seedling establishment in these soils is favored in species with
larger seeds with greater nutrient levels (Stock
et al.
1990
; Jurado & Westoby
1992
; Bond
et al.
1999
) and this makes them a target for seed predators already
stressed by the oligotrophic conditions in this ecosystem. This pressure is even
greater considering that the high nutrient stores in seeds will limit seed number
over that compared with less nutrient stressed soils. In general, these costs are
hard to recoup over the higher production of smaller seeds (Moles
et al.
2004
)
typical of species with soil-stored seedbanks, suggesting there are other rewards to
canopy storage.
It is hypothesized that interactions between selection for large seeds and avoid-
ance of seed predation have been important drivers in the evolution of serotiny in
these oligotrophic environments (
Fig. 9.7
). Serotiny puts significant amounts of
carbon, which is more readily available than other nutrients, into protecting the
seeds against predispersal predation and since total seedbank size is generally